Upload presentasi
Presentasi sedang didownload. Silahkan tunggu

1
TERBENTUKNYA TURUNAN “BARU”
ALAMIAH PERBAIKAN GENERASI 1. KAWIN SILANG - SEKSUAL PERBIAKAN UNGGULAN TURUNAN BARU POPULASI MIKROBA RESISTEN KONYUGASI 2. PARASEKSUAL TRANSFORMASI PERPINDAHAN SIFAT RESISTENSI MIKROBA TERHADAP ANTIBIOTIK TRANSDUKSI EVOLUSI SINAR MATAHARI-KOSMIS POLUTAN / CEMARAN BIBIT UNGGUL DARI BIJI TOKSISITAS OBAT SPONTAN 3. MUTASI KIMIA SINAR PENGION ANTIBODI MONOKLONAL 4. FUSI SEL HIBRIDA SOMATIK : TANAMAN UNGGULAN TURUNAN DENGAN SIFAT BAIK KEDUANYA CYBRIDE

2
TERBENTUKNYA TURUNAN BARU
PRODUKSI PROTEIN BIOAKTIF PADA ORGANISME LAIN ( INSULIN, VAKSIN-HB ) 5. REKAYASA GENETIK PROTEIN ENGINEERING 6. TRANSGENIK PERUBAHAN GEN --> FENOTIP DAN PRODUK BARU TERBENTUKNYA TURUNAN IDENTIK VEGETATIP MEMPERBANYAK DIRI IDENTIK ALAMIAH TURUNAN UNGGULAN YANG SAMA UNGGULAN PRODUK BARU KLONING ORGANISME
 5. REKAYASA GENETIK. PROTEIN ENGINEERING. 6. TRANSGENIK. PERUBAHAN GEN --> FENOTIP DAN PRODUK BARU. TERBENTUKNYA TURUNAN IDENTIK. VEGETATIP. MEMPERBANYAK DIRI IDENTIK ALAMIAH. TURUNAN UNGGULAN YANG SAMA. UNGGULAN PRODUK BARU. KLONING ORGANISME.")
3
RESISTENSI MIKROBA PATOGEN TERHADAP ANTIBIOTIK Bakteri Virus Parasit

4
Bacterial cells divide by the process of binary fission
Bacterial cells divide by the process of binary fission. A cell will first duplicate its chromosome, elongate, and then pinch in the middle creating two genetically identical daughter cells. During replication of the chromosome, there is no built in repair mechanism, so if mistakes occur during replication they will be passed on to daughter cells. Some mistakes have no effect on the bacterial cell. Other mistakes can create fatal errors and the cell will die. However, some mistakes provide new variants genes.

12
Bacterial cells divide by the process of binary fission
Bacterial cells divide by the process of binary fission. A cell will first duplicate its chromosome, elongate, and then pinch in the middle creating two genetically identical daughter cells. During replication of the chromosome, there is no built in repair mechanism, so if mistakes occur during replication they will be passed on to daughter cells. Some mistakes have no effect on the bacterial cell. Other mistakes can create fatal errors and the cell will die. However, some mistakes provide new variants genes.

13
HOW - WHY ? ANTIBIOTIC-RESISTANT BACTERIA 1. GENE FOR “EFFLUX” PUMPS
THAT EJECT ANTIBIOTICS FROM CELL 2. GEN FOR ANTIBIOTIC ALTERING ENZYME/TARGET 3. GEN FOR ANTIBIOTIC DEGRADING ENZYME OTHER GENETIC MECHANISM ???

14
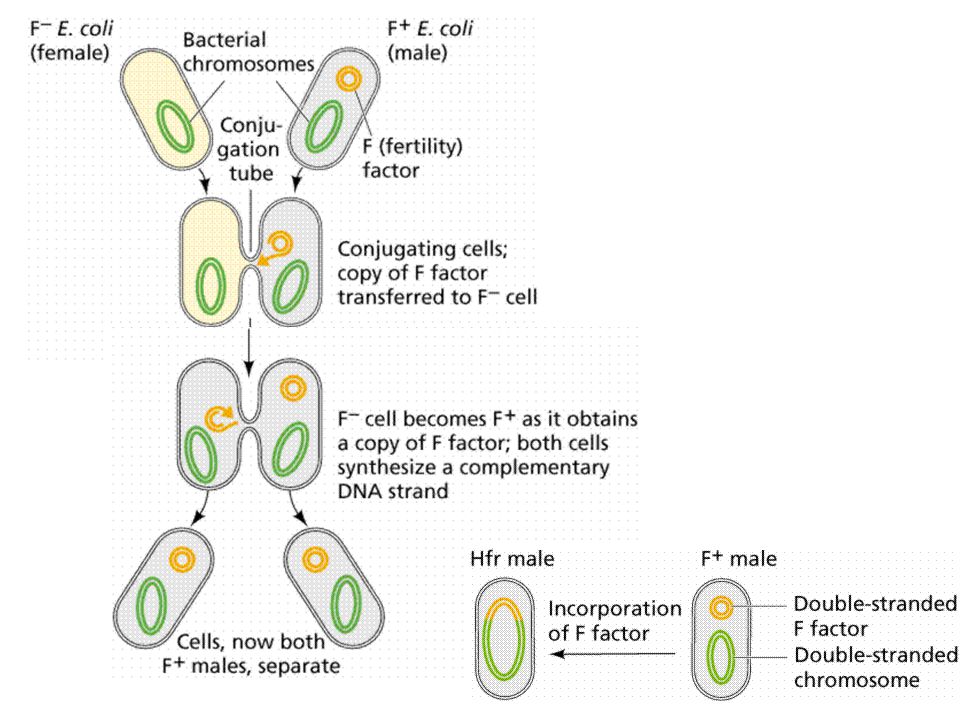
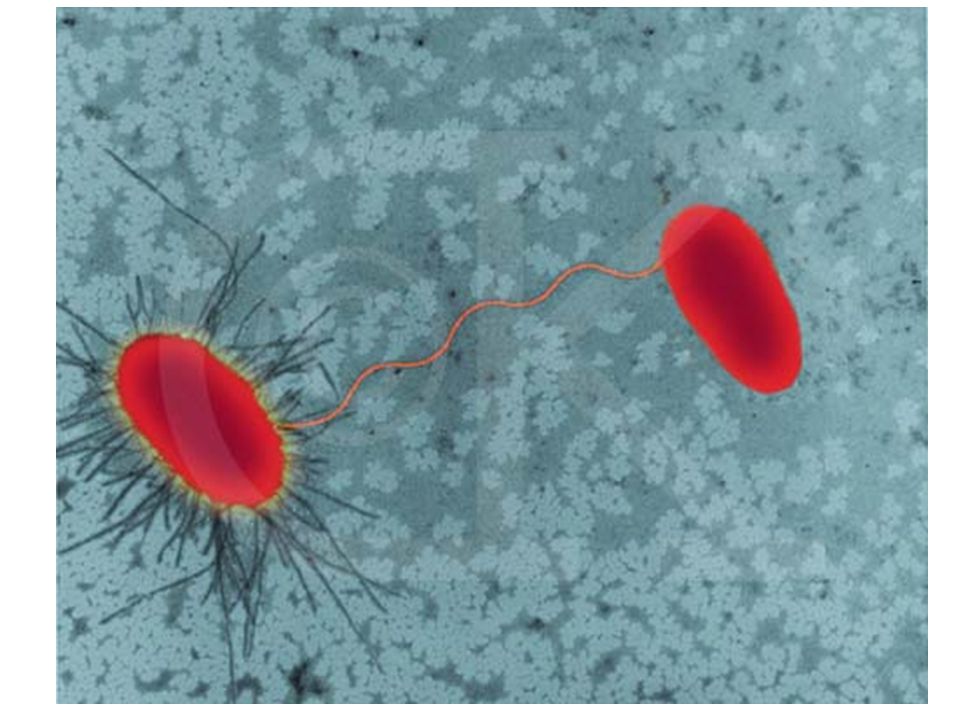
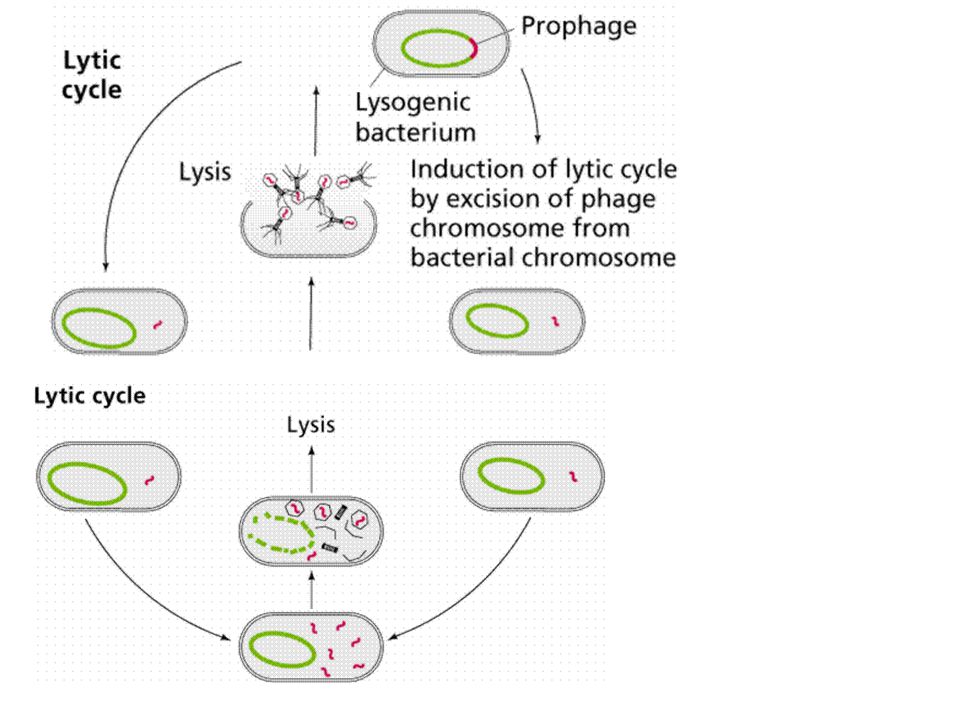
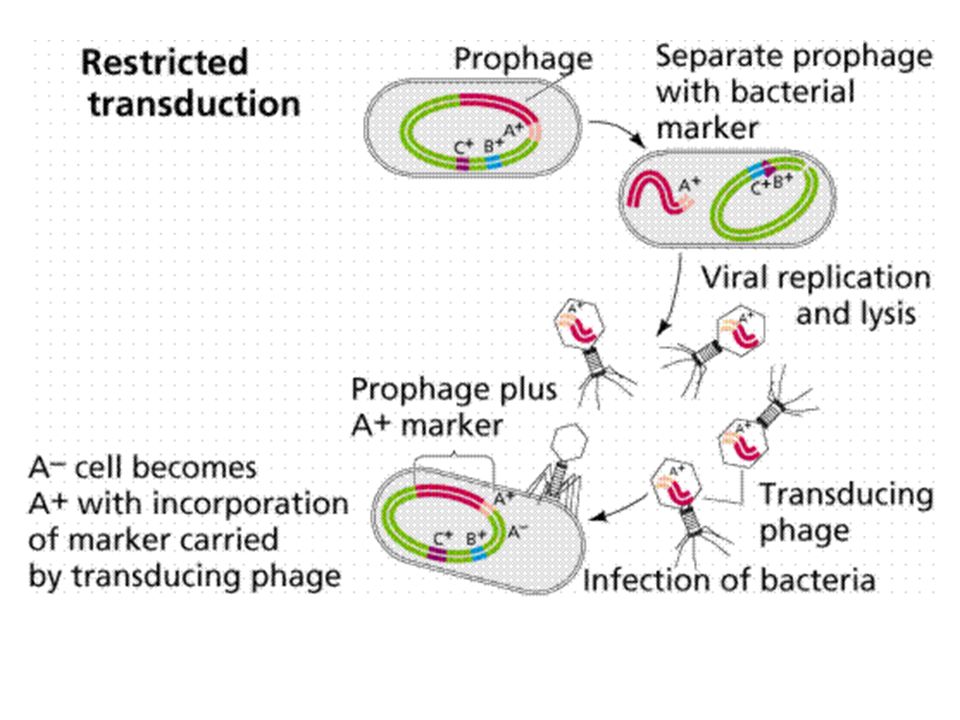
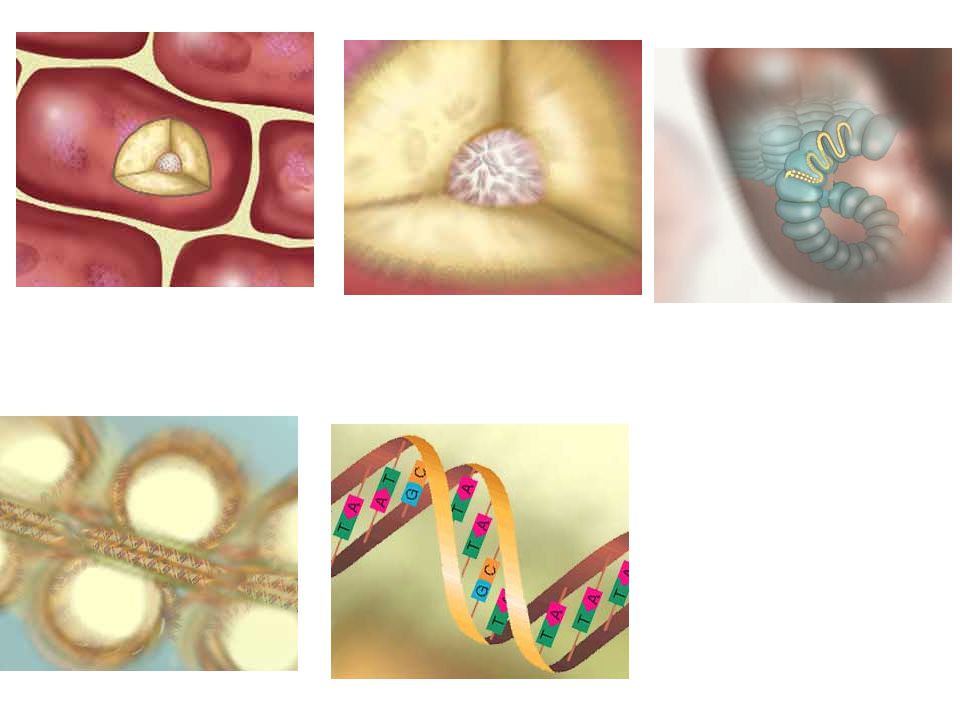
BACTERIA PICK UP RESISTANCE GENES KONYUGASI TRANSFORMASI TRANSDUKSI

15
sensitive bacterium resistance bacterium DNA-frament = resistance gene transformation sensitive bacterium

16
conjugation

17
transduction

18
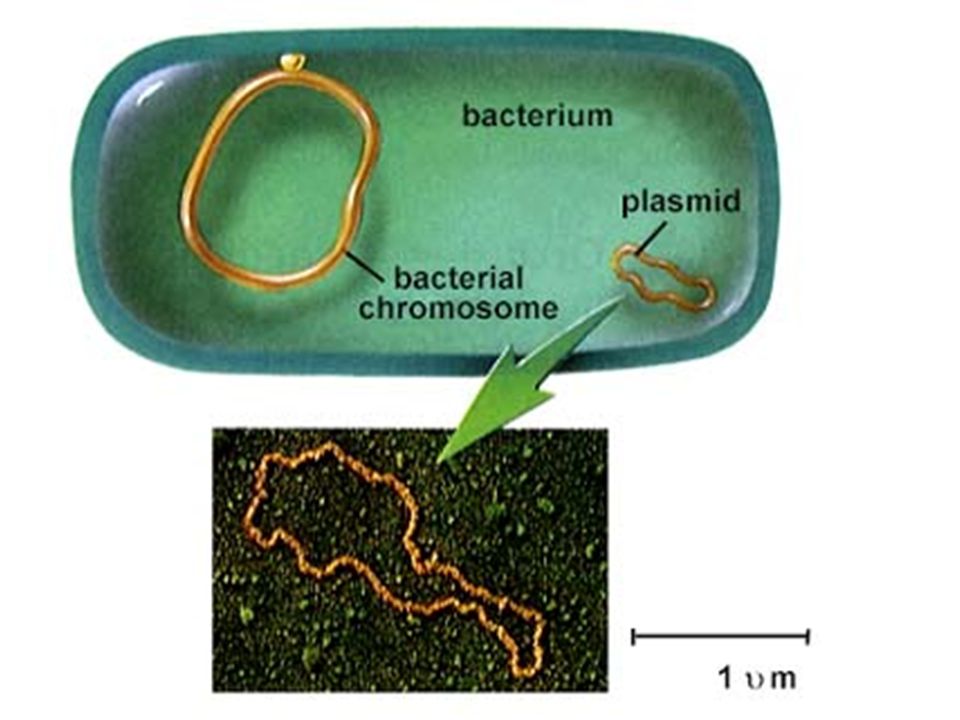
ANTIBIOTIC-RESISTANT BACTERIA owe their drug insensitivity to resistance genes. For example, such genes might code for "efflux" pumps that eject antibiotics from cells (a). Or the genes might give rise to enzymes that degrade the antibiotics (b) or that chemically alter--and inactivate--the drugs (c). Resistance genes can reside on the bacterial chromosome or, more typically, on small rings of DNA called plasmids. Some of the genes are inherited, some emerge through random mutations in bacterial DNA, and some are imported from other bacteria. BACTERIA PICK UP RESISTANCE GENES from other bacterial cells in three main ways. Often they receive whole plasmids bearing one or more such genes from a donor cell (a). Other times, a virus will pick up a resistance gene from one bacterium and inject it into a different bacterial cell (b). Alternatively, bacteria sometimes scavenge gene-bearing snippets of DNA from dead cells in their vicinity (c). Genes obtained through viruses or from dead cells persist in their new owner if they become incorporated stably into the recipient's chromosome or into a plasmid.
. Or the genes might give rise to enzymes that degrade the antibiotics (b) or. that chemically alter--and inactivate--the drugs (c). Resistance genes can reside on the bacterial chromosome or, more typically, on small rings of DNA called plasmids. Some of the genes are inherited, some emerge through random mutations in bacterial. DNA, and some are imported from other bacteria. BACTERIA PICK UP RESISTANCE GENES from other bacterial cells in three main ways. Often they receive whole plasmids bearing one or more such genes from a donor cell (a). Other times, a virus will pick up a resistance gene from one. bacterium and inject it into a different bacterial cell (b). Alternatively, bacteria sometimes scavenge gene-bearing snippets of DNA from dead cells in their vicinity (c). Genes obtained through viruses or from dead cells persist in their new owner if they become incorporated stably into the recipient s chromosome or into a plasmid.")
19
Genetic Basis of Acquired Resistance
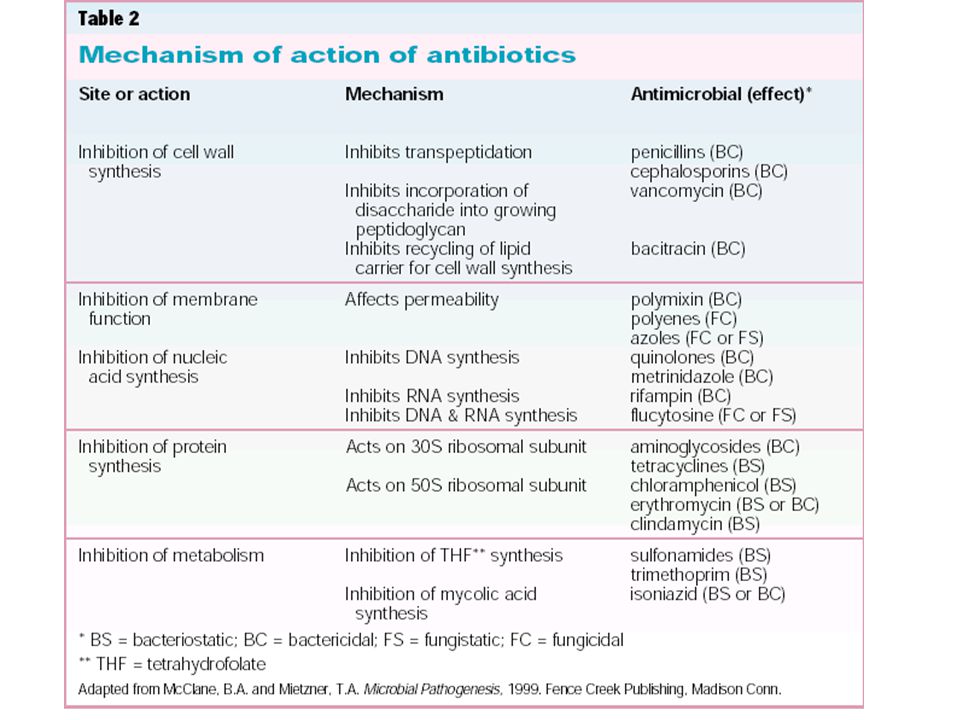
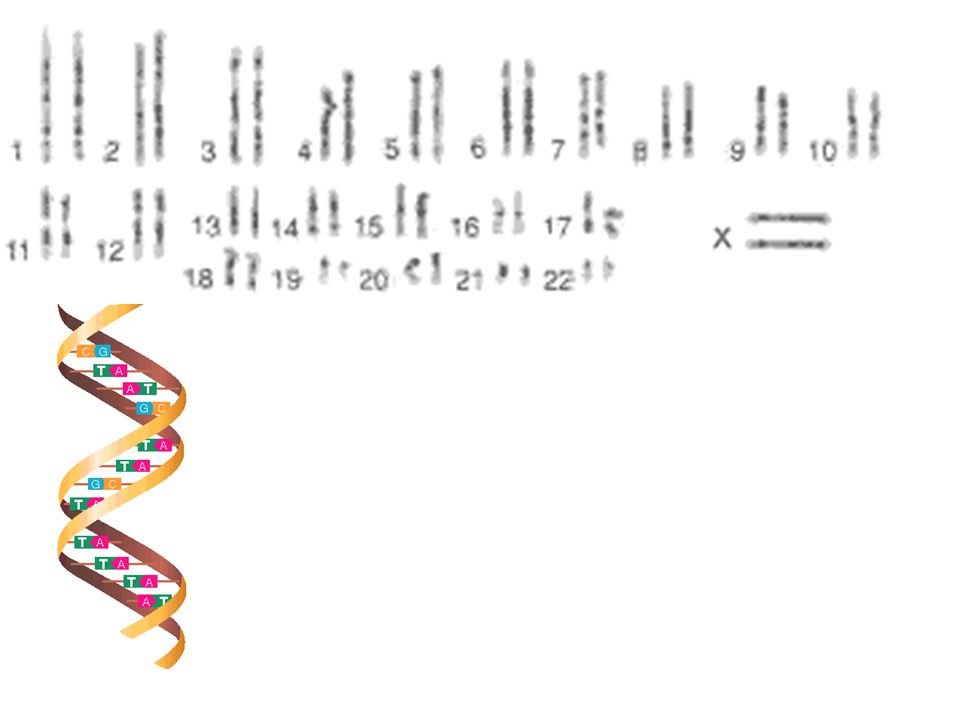
Mutation - spontaneous alteration in genetic sequence in bacterial chromosome Conjugation - transfer of genetic information via conjugation tubes Transduction (bacteriophage) - phage incorporation of genetic information which can then be transferred to another bacterium Transformation - possible assimilation of genetic material across cell wall/membrane Location: plasmids, transposons, chromosome Phenotypic Mechanisms of Resistance Enzyme degradation Barrier to the target Target absent
 - phage incorporation of genetic information which can then be transferred to another bacterium. Transformation - possible assimilation of genetic material across cell wall/membrane. Location: plasmids, transposons, chromosome. Phenotypic Mechanisms of Resistance. Enzyme degradation. Barrier to the target. Target absent.")
20
Antibiotic resistance results from gene action
Antibiotic resistance results from gene action. Bacteria acquire genes conferring resistance in any of three ways In spontaneous DNA mutation, bacterial DNA (genetic material) may mutate (change) spontaneously (indicated by starburst). Drug- resistant tuberculosis arises this way. In a form of microbial sex called transformation, one bacterium may take up DNA from another bacterium. Pencillin-resistant gonorrhea results from transformation Most frightening, however, is resistance acquired from a small circle of DNA called a plasmid, that can flit from one type of bacterium to another. A single plasmid can provide a slew of different resistances
 may mutate (change) spontaneously (indicated by starburst). Drug- resistant tuberculosis arises this way. In a form of microbial sex called transformation, one bacterium may take up DNA from another bacterium. Pencillin-resistant gonorrhea results from transformation. Most frightening, however, is resistance acquired from a small circle of DNA called a plasmid, that can flit from one type of bacterium to another. A single plasmid can provide a slew of different resistances.")
30
pump blocker

31
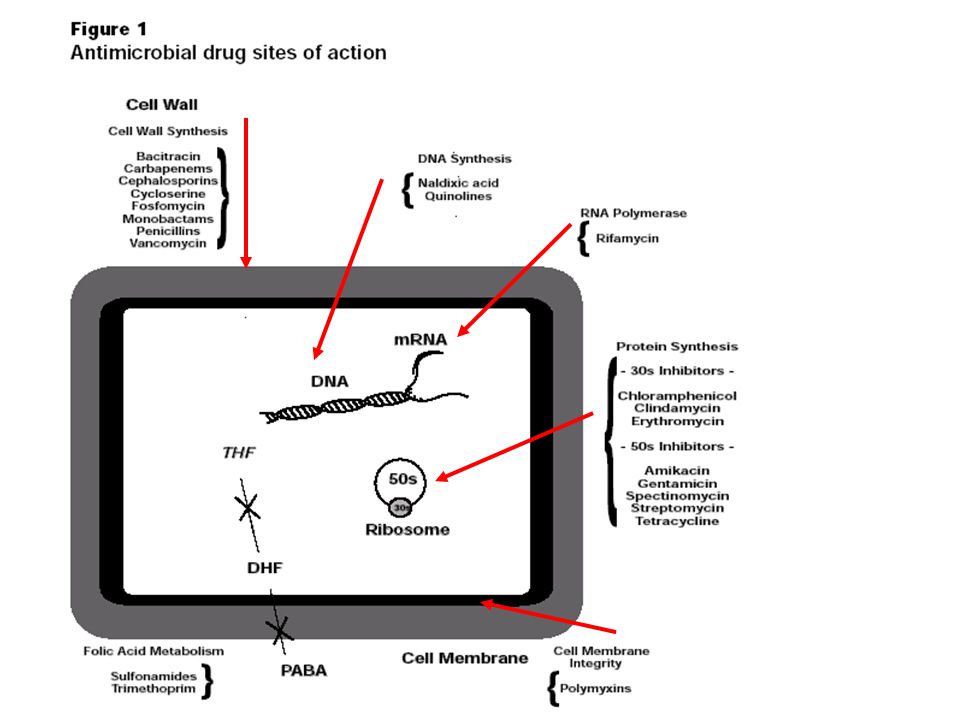
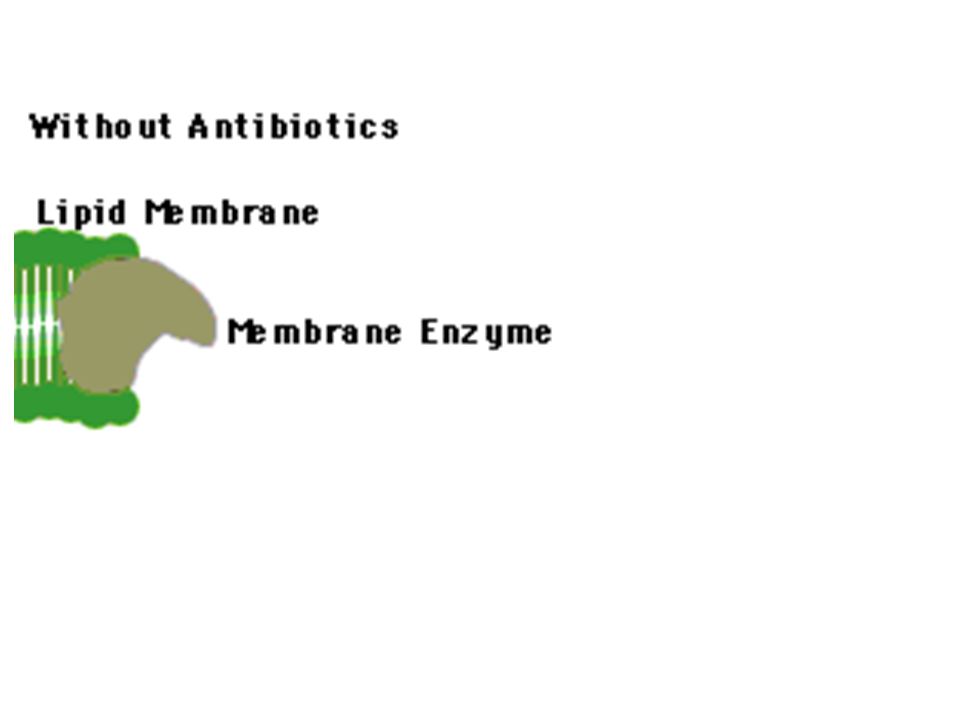
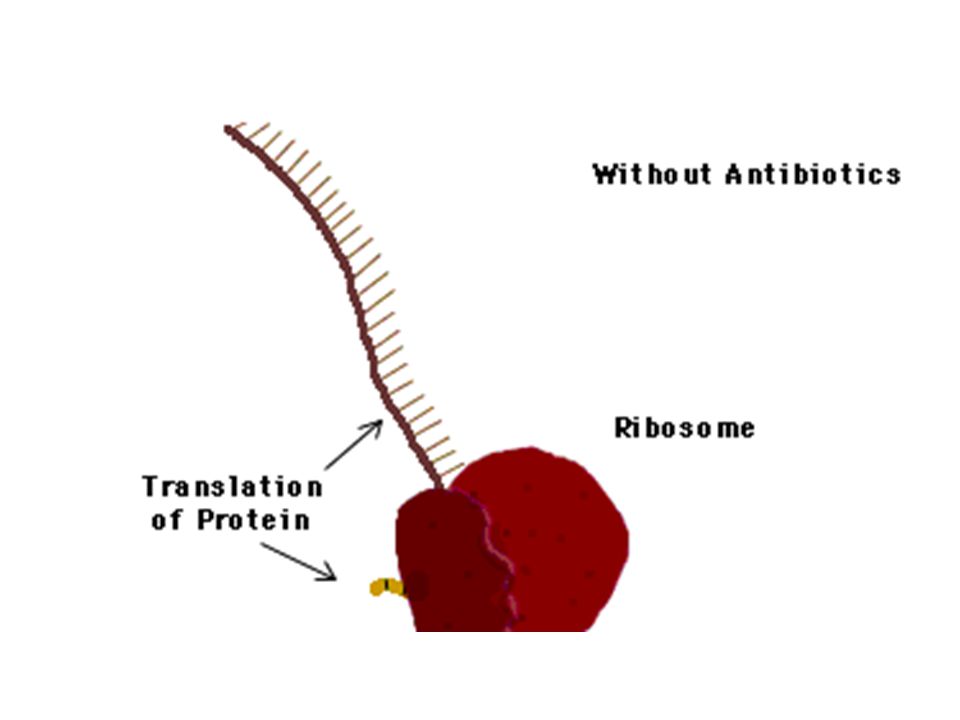
ONE PHARMACEUTICAL STRATEGY for overcoming resistance capitalizes on the discovery that some bacteria defeat certain antibiotics, such as tetracycline, by pumping out the drugs (a). To combat that ploy, investigators are devising compounds that would jam the pumps (b), thereby freeing the antibiotics to function effectively. In the case of tetracycline, the antibiotic works by interfering with the ribosomes that manufacture bacterial proteins.
, thereby freeing the antibiotics to function effectively. In the case of tetracycline, the antibiotic works by interfering with the ribosomes that manufacture bacterial proteins..")
32
RESISTANT POPULATION of bacteria will disappear naturally only if susceptible bacteria live in the vicinity. After antibiotic therapy stops (a), resistant bacteria can persist for a while. If susceptible bacteria are nearby, however, they may recolonize the individual (b). In the absence of the drug, the susceptible bugs will have a slight survival advantage because they do not have to expend energy maintaining resistance genes. After a time, then, they may outcompete the resistant microbes (c and d). For this reason, protecting susceptible bacteria needs to be a public health priority.
, resistant bacteria can persist for a while. If susceptible bacteria are nearby, however, they may recolonize the individual (b). In the absence of the drug, the susceptible bugs will have a slight survival advantage because they do not have to expend energy maintaining resistance genes. After a time, then, they may outcompete the resistant microbes (c and d). For this reason, protecting susceptible bacteria needs to be a public health priority..")
33
Resistensi bakteri terhadap suatu antibiotika terkait adanya suatu gen
Resistensi bakteri terhadap suatu antibiotika terkait adanya suatu gen. Jelaskan adanya 3 jenis gen yang kalau diekspresikan dapat menjadi dasar mekanisme resistensi bakteri tsb.), Jelaskan (gambar dan uraian). Resistensi mikroba terhadap suatu antibiotika dapat disebabkan karena terjadinya mutasi gen ( yang spontan, adaptasi) atau mikroba memperoleh gen resisten dari mikroba lainnya (terbentuknya turunan baru/strain/mutan secara paraseksual/ parameiotik). Jelaskan bagaimana 3 proses paraseksual tersebut.(gambar dan uraian) STOP
, Jelaskan (gambar dan uraian). Resistensi mikroba terhadap suatu antibiotika dapat disebabkan karena terjadinya mutasi gen ( yang spontan, adaptasi) atau mikroba memperoleh gen resisten dari mikroba lainnya (terbentuknya turunan baru/strain/mutan secara paraseksual/ parameiotik). Jelaskan bagaimana 3 proses paraseksual tersebut.(gambar dan uraian) STOP.")
46
ADVANCE

49
IMMUNOLOGY

50
Figure 1. Function of Interdigitating Dendritic Cells
Figure 1. Function of Interdigitating Dendritic Cells. Pathogen-associated molecular patterns (PAMPs) allow the pattern-recognition receptors on the dendritic cells and macrophages of the innate immune response to differentiate between potentially harmful foreign microorganisms and self constituents. These cells are also stimulated by endogenous activators such as interferon- , heat-shock proteins, and tumor necrosis factor that are released as a result of infection. The activated antigen-presenting cells then present a cell-surface complex of a major-histocompatibility-complex (MHC) molecule and peptide, derived by intracellular processing of the foreign antigen, to the T-cell receptors on the highly specific CD28-bearing naive T cells, which become activated in the acquired immune response. Activation also causes dendritic cells to enhance their expression of B7 costimulatory molecules.
 allow the pattern-recognition receptors on the dendritic cells and macrophages of the innate immune response to differentiate between potentially harmful foreign microorganisms and self constituents. These cells are also stimulated by endogenous activators such as interferon- , heat-shock proteins, and tumor necrosis factor that are released as a result of infection. The activated antigen-presenting cells then present a cell-surface complex of a major-histocompatibility-complex (MHC) molecule and peptide, derived by intracellular processing of the foreign antigen, to the T-cell receptors on the highly specific CD28-bearing naive T cells, which become activated in the acquired immune response. Activation also causes dendritic cells to enhance their expression of B7 costimulatory molecules.")
51
Figure 2. A System Used by Natural Killer Cells to Recognize Normal Cells and Cells That Lack Major-Histocompatibility-Complex Class I Surface Molecules. Killer-activating receptors recognize a number of molecules present on the surface of normal, nucleated cells, and in the absence of an inhibitory signal from killer-inhibitory receptors, which recognize major-histocompatibility-complex (MHC) class I molecules, the receptors issue an order to the natural killer cells to attack and kill the other cell. The cytotoxic granules of the natural killer cells, which contain perforin and granzymes, become polarized at the interface with the target cell and are then released into the cell.
 class I molecules, the receptors issue an order to the natural killer cells to attack and kill the other cell. The cytotoxic granules of the natural killer cells, which contain perforin and granzymes, become polarized at the interface with the target cell and are then released into the cell..")
52
Figure 3. The Acute Inflammatory Response
Figure 3. The Acute Inflammatory Response. Neutrophils are among the first cells to arrive at the scene of an infection and are important contributors to the acute inflammatory response. As the neutrophil rolls along the blood-vessel wall, the L-selectin on its surface binds to carbohydrate structures such as sialyl-Lewisx on the adhesion molecules on the vascular endothelium, and its progress is eventually halted. As the neutrophil becomes activated, it replaces L-selectin with other cell-surface adhesion molecules, such as integrins. These molecules bind E-selectin, which is present on the blood-vessel wall as a result of the influence of inflammatory mediators such as bacterial lipopolysaccharides and the cytokines interleukin-1 and tumor necrosis factor . The activated neutrophil then enters the tissues, where it is attracted to the infection site by a number of chemoattractants. The neutrophil can then phagocytose and destroy the C3b-coated bacteria.

53
Figure 4. Structure of Immature and Mature B-Cell and T-Cell Antigen Receptors. The immature pre–B cells and pre–T cells express preliminary versions of the antigen receptor. At this stage, the B-cell receptor comprises a pair of heavy (H) chains, each with a variable (V) and a constant (Cµ) region identical to those found in the mature receptor, and a pair of surrogate light chains, termed Vpre-B and 5. As the B cell develops, the surrogate light chains are replaced by conventional light (L) chains of either the or type, each with a variable and a constant region. This mature IgM molecule acts as the B-cell receptor for antigen, usually together with IgD B-cell receptors with the same antigen specificity. The variable regions of the heavy and light chains each contain three hypervariable complementarity-determining regions (CDRs). The CDRs make contact with the antigen. One of the two antigen-binding arms (Fab) of the bivalent antibody molecule is indicated. The circulating version of the antibody contains the same four chains but lacks the transmembrane sequence that anchors the B-cell receptor in the lymphocyte membrane. With respect to T cells, the immature T-cell receptor consists of a ß chain identical to that found in the mature receptor and a pre-T chain that comprises only a constant region. This segment is replaced by an chain to form the mature T-cell receptor, and each chain consists of a variable region and a constant region. For the sake of simplicity, the antigen receptors are shown without their associated signal-transduction units.
 chains, each with a variable (V) and a constant (Cµ) region identical to those found in the mature receptor, and a pair of surrogate light chains, termed Vpre-B and 5. As the B cell develops, the surrogate light chains are replaced by conventional light (L) chains of either the or type, each with a variable and a constant region. This mature IgM molecule acts as the B-cell receptor for antigen, usually together with IgD B-cell receptors with the same antigen specificity. The variable regions of the heavy and light chains each contain three hypervariable complementarity-determining regions (CDRs). The CDRs make contact with the antigen. One of the two antigen-binding arms (Fab) of the bivalent antibody molecule is indicated. The circulating version of the antibody contains the same four chains but lacks the transmembrane sequence that anchors the B-cell receptor in the lymphocyte membrane. With respect to T cells, the immature T-cell receptor consists of a ß chain identical to that found in the mature receptor and a pre-T chain that comprises only a constant region. This segment is replaced by an chain to form the mature T-cell receptor, and each chain consists of a variable region and a constant region. For the sake of simplicity, the antigen receptors are shown without their associated signal-transduction units..")
54
Figure 5. Diversity of Antigen Receptors
Figure 5. Diversity of Antigen Receptors. The enormously diverse specificities of the antigen receptors are produced by gene rearrangements during the early developmental stages of the lymphocyte. The events involved in generating a coding sequence for the immunoglobulin heavy chain are shown. Early in B-cell development, pro–B cells mature into pre–B cells, at which stages they express the recombination-activating genes RAG1 and RAG2. The recombinases encoded by these genes mediate the random rearrangement of 1 of 25 diversity (D ) gene segments next to any 1 of 6 joining (J ) gene segments. This is followed by the rearrangement of any 1 of 50 variable (V ) gene segments next to the already rearranged DJ segment. Different B cells will rearrange a different segment in each pool, thereby creating one level of diversity. Further diversity is brought about by splicing inaccuracies and by the incorporation of nucleotides mediated by the enzyme terminal deoxyribonucleotidyltransferase (TdT). The heavy-chain primary RNA transcript is processed into messenger RNA (mRNA), with splicing of the rearranged VDJ segment next to the constant (C ) region gene. This mRNA will encode a heavy chain that appears on the surface of the pre–B cell together with the surrogate light chain, which is encoded by genes that do not undergo rearrangement. As the pre–B cell continues to mature, the immunoglobulin light-chain genes undergo rearrangement; the resulting light chain replaces the surrogate light chain, and thereby produces a mature IgM B-cell receptor on the cell surface. The B-cell receptors at this stage also usually include IgD antibodies with the same specificity as the IgM molecule, produced by alternative splicing of the rearranged VDJ to either the Cµ or the C gene. The expression of RAG1 and RAG2 is then switched off. After encountering an antigen, and in the presence of costimulatory signals, the B cell further differentiates into a plasma cell, which secretes high levels of the specific antibody (or into a memory B cell). The same general principles regarding the rearrangement process apply to the generation of /ß and / T-cell receptors. The gene segments in the figure are not drawn to scale.
 gene segments next to any 1 of 6 joining (J ) gene segments. This is followed by the rearrangement of any 1 of 50 variable (V ) gene segments next to the already rearranged DJ segment. Different B cells will rearrange a different segment in each pool, thereby creating one level of diversity. Further diversity is brought about by splicing inaccuracies and by the incorporation of nucleotides mediated by the enzyme terminal deoxyribonucleotidyltransferase (TdT). The heavy-chain primary RNA transcript is processed into messenger RNA (mRNA), with splicing of the rearranged VDJ segment next to the constant (C ) region gene. This mRNA will encode a heavy chain that appears on the surface of the pre–B cell together with the surrogate light chain, which is encoded by genes that do not undergo rearrangement. As the pre–B cell continues to mature, the immunoglobulin light-chain genes undergo rearrangement; the resulting light chain replaces the surrogate light chain, and thereby produces a mature IgM B-cell receptor on the cell surface. The B-cell receptors at this stage also usually include IgD antibodies with the same specificity as the IgM molecule, produced by alternative splicing of the rearranged VDJ to either the Cµ or the C gene. The expression of RAG1 and RAG2 is then switched off. After encountering an antigen, and in the presence of costimulatory signals, the B cell further differentiates into a plasma cell, which secretes high levels of the specific antibody (or into a memory B cell). The same general principles regarding the rearrangement process apply to the generation of /ß and / T-cell receptors. The gene segments in the figure are not drawn to scale.")
55
Figure 6. Recognition of Epitopes by B Cells
Figure 6. Recognition of Epitopes by B Cells. Using the antibody molecule as its receptor, the B cell recognizes epitopes on the surface of the antigen. If it is stimulated by this contact, the B cell proliferates, and the resulting clones can secrete antibody whose specificity is the same as that of the cell-surface receptor that bound the epitope. Responses usually involve several different clones of lymphocytes and are therefore referred to as polyclonal. Although not shown here, for each epitope there may be several different lymphocyte clones with different B-cell receptors, each of which recognizes the epitope in a slightly different way and therefore with a different binding strength (affinity).
.")
56
Figure 7. Positive and Negative Selection in the Thymus
Figure 7. Positive and Negative Selection in the Thymus. T cells need to detect foreign antigens presented by self major-histocompatibility-complex (MHC) molecules. Part of the T-cell receptor recognizes the foreign peptide, and part of it recognizes the self MHC molecule. The random nature of T-cell–receptor gene rearrangements means that only a minority of T cells are capable of performing this task. Many of the immature CD4 and CD8 double-positive T cells are useless because their T-cell receptors do not recognize self MHC molecules at all. These T cells eventually undergo apoptosis. Cells whose T-cell receptors have various affinities for binding self MHC molecules (usually containing a self peptide) are positively selected on cortical epithelial cells. However, many of these cells are potentially harmful because their T-cell receptors have a high affinity for a complex of self peptide and a self MHC molecule (or even an MHC molecule alone). These autoimmune T cells are eliminated by the induction of apoptosis when they interact with dendritic cells and macrophages in the thymic medulla (negative selection ). This leaves T cells with only a weak affinity for self MHC molecules. These cells form the pool of T cells that are exported from the thymus as single-positive (CD4 or CD8) cells. In the periphery they have the potential to recognize a complex of foreign peptide plus self MHC molecules and to become activated if the affinity of the interaction exceeds a certain threshold.
 molecules. Part of the T-cell receptor recognizes the foreign peptide, and part of it recognizes the self MHC molecule. The random nature of T-cell–receptor gene rearrangements means that only a minority of T cells are capable of performing this task. Many of the immature CD4 and CD8 double-positive T cells are useless because their T-cell receptors do not recognize self MHC molecules at all. These T cells eventually undergo apoptosis. Cells whose T-cell receptors have various affinities for binding self MHC molecules (usually containing a self peptide) are positively selected on cortical epithelial cells. However, many of these cells are potentially harmful because their T-cell receptors have a high affinity for a complex of self peptide and a self MHC molecule (or even an MHC molecule alone). These autoimmune T cells are eliminated by the induction of apoptosis when they interact with dendritic cells and macrophages in the thymic medulla (negative selection ). This leaves T cells with only a weak affinity for self MHC molecules. These cells form the pool of T cells that are exported from the thymus as single-positive (CD4 or CD8) cells. In the periphery they have the potential to recognize a complex of foreign peptide plus self MHC molecules and to become activated if the affinity of the interaction exceeds a certain threshold.")
57
Figure 8. The Germinal Center
Figure 8. The Germinal Center. During the initiation of the acquired immune response, germinal centers form in the secondary lymphoid tissues in order to create a microenvironment where all the necessary antigen-specific and innate antigen-presenting cells can interact. Several cytokines, such as interleukin-2, 4, 6, and 10 and transforming growth factor ß, and various cell-surface molecules, including CD40, CD19, CD21, and B7, are critically important for these interactions. Antigen-stimulated proliferation of B cells occurs in the dark zone and is accompanied by the fine-tuning of specificity resulting from somatic hypermutation of the immunoglobulin variable-region genes. On reaching the basal light zone, high-affinity antigen-specific B cells are positively selected as a result of their interaction with antigen–antibody complexes on the surface of follicular dendritic cells. B cells that are not positively selected undergo apoptosis and are phagocytosed by tingible-body macrophages. The positively selected cells migrate to the apical light zone, where proliferation continues, class switching occurs, and memory cells and plasma-cell precursors are generated.

58
Figure 9. Activation of T Cells
Figure 9. Activation of T Cells. The activation of T cells involves a highly complex series of integrated events that result from the cross-linking of the antigen receptor on the cell surface. Because the antigen receptors have extremely short cytoplasmic tails, they are associated (in T cells) with the CD3 and chain signal-transduction molecules bearing cytoplasmic immunoreceptor tyrosine-based activation motifs (ITAMs), which are subject to phosphorylation (P) by protein kinases such as p56lck, p59fyn, and ZAP-70 (for simplicity, only one of the CD3 molecules is shown). The initial stages of activation also involve the binding of p56lck to the cytoplasmic tail of CD4 (in helper T cells) or CD8 (in cytotoxic T cells). These events lead to downstream signaling involving a number of different biochemical pathways and ultimately to the transcriptional activation of genes involved in cellular proliferation and differentiation. Signals from costimulatory receptors such as CD28 and CD154 must also be present in order to activate the lymphocyte; in the event that signals are sent only from the antigen-receptor signal-transducing molecules, anergy or apoptosis will occur.
 with the CD3 and chain signal-transduction molecules bearing cytoplasmic immunoreceptor tyrosine-based activation motifs (ITAMs), which are subject to phosphorylation (P) by protein kinases such as p56lck, p59fyn, and ZAP-70 (for simplicity, only one of the CD3 molecules is shown). The initial stages of activation also involve the binding of p56lck to the cytoplasmic tail of CD4 (in helper T cells) or CD8 (in cytotoxic T cells). These events lead to downstream signaling involving a number of different biochemical pathways and ultimately to the transcriptional activation of genes involved in cellular proliferation and differentiation. Signals from costimulatory receptors such as CD28 and CD154 must also be present in order to activate the lymphocyte; in the event that signals are sent only from the antigen-receptor signal-transducing molecules, anergy or apoptosis will occur.")
59
Figure 10. An Overview of Lymphocyte Responses
Figure 10. An Overview of Lymphocyte Responses. T cells characteristically possess T-cell receptors (TCRs) that recognize processed antigen presented by major-histocompatibility-complex (MHC) molecules, as shown on the left-hand side of the figure. Most cytotoxic T cells are positive for CD8, recognize processed antigen presented by MHC class I molecules, and kill infected cells, thereby preventing viral replication. Activated cytotoxic T cells secrete interferon- that, together with interferon- and interferon-ß produced by the infected cells themselves, sets up a state of cellular resistance to viral infection. As shown on the right-hand side of the figure, helper T cells are generally positive for CD4, recognize processed antigen presented by MHC class II molecules, and can be divided into two major populations. Type 1 (Th1) helper T cells secrete interferon- and interleukin-2, which activate macrophages and cytotoxic T cells to kill intracellular organisms; type 2 (Th2) helper T cells secrete interleukin-4, 5, and 6, which help B cells secrete protective antibodies. B cells recognize antigen either directly or in the form of immune complexes on follicular dendritic cells in germinal centers.
 that recognize processed antigen presented by major-histocompatibility-complex (MHC) molecules, as shown on the left-hand side of the figure. Most cytotoxic T cells are positive for CD8, recognize processed antigen presented by MHC class I molecules, and kill infected cells, thereby preventing viral replication. Activated cytotoxic T cells secrete interferon- that, together with interferon- and interferon-ß produced by the infected cells themselves, sets up a state of cellular resistance to viral infection. As shown on the right-hand side of the figure, helper T cells are generally positive for CD4, recognize processed antigen presented by MHC class II molecules, and can be divided into two major populations. Type 1 (Th1) helper T cells secrete interferon- and interleukin-2, which activate macrophages and cytotoxic T cells to kill intracellular organisms; type 2 (Th2) helper T cells secrete interleukin-4, 5, and 6, which help B cells secrete protective antibodies. B cells recognize antigen either directly or in the form of immune complexes on follicular dendritic cells in germinal centers.")
60
Figure 11. Role of Antibodies. Antibodies rarely act in isolation
Figure 11. Role of Antibodies. Antibodies rarely act in isolation. Their usual role is to focus components of the innate immune system on the pathogen, and the activation of these destructive forces normally requires coordinating events that occur after Fab heavy- and light-chain variable regions (VH and VL) of the antibody are bound to antigen, leading to the display of multiple exposed Fc regions. The figure shows two examples of this process: the activation of the classic complement pathway after binding of C1q to Fc, and the activation of phagocytosis after the cross-linking of Fc receptors and binding of the Fc R on the macrophage.
 of the antibody are bound to antigen, leading to the display of multiple exposed Fc regions. The figure shows two examples of this process: the activation of the classic complement pathway after binding of C1q to Fc, and the activation of phagocytosis after the cross-linking of Fc receptors and binding of the Fc R on the macrophage.")
61
complement

62
Figure 1. The Three Activation Pathways of Complement: the Classical, Mannose-Binding Lectin, and Alternative Pathways. The three pathways converge at the point of cleavage of C3. The classical pathway is initiated by the binding of the C1 complex (which consists of C1q, two molecules of C1r, and two molecules of C1s) to antibodies bound to an antigen on the surface of a bacterial cell. C1s first cleaves C4, which binds covalently to the bacterial surface, and then cleaves C2, leading to the formation of a C4b2a enzyme complex, the C3 convertase of the classical pathway. The mannose-binding lectin pathway is initiated by binding of the complex of mannose-binding lectin and the serine proteases mannose-binding lectin–associated proteases 1 and 2 (MASP1 and MASP2, respectively) to arrays of mannose groups on the surface of a bacterial cell. MASP2 acts in a fashion similar to that of C1s to lead to the formation of the C3 convertase enzyme C4b2a. MASP1 may be able to cleave C3 directly. The alternative pathway is initiated by the covalent binding of a small amount of C3b to hydroxyl groups on cell-surface carbohydrates and proteins and is activated by low-grade cleavage of C3 in plasma. This C3b binds factor B, a protein homologous to C2, to form a C3bB complex. Factor D cleaves factor B bound to C3b to form the alternative pathway C3 complex C3bBb. The binding of properdin stabilizes this enzyme. The C3 convertase enzymes cleave many molecules of C3 to C3b, which bind covalently around the site of complement activation. Some of this C3b binds to the C4b and C3b in the convertase enzymes of the classical and alternative pathways, respectively, forming C5 convertase enzymes. This C3b acts as an acceptor site for C5, which is cleaved to form the anaphylatoxin C5a and C5b, which initiates the formation of the membrane-attack complex. The activities of biologically active proteins and protein fragments of the complement pathway are described in (Table 1).
 to antibodies bound to an antigen on the surface of a bacterial cell. C1s first cleaves C4, which binds covalently to the bacterial surface, and then cleaves C2, leading to the formation of a C4b2a enzyme complex, the C3 convertase of the classical pathway. The mannose-binding lectin pathway is initiated by binding of the complex of mannose-binding lectin and the serine proteases mannose-binding lectin–associated proteases 1 and 2 (MASP1 and MASP2, respectively) to arrays of mannose groups on the surface of a bacterial cell. MASP2 acts in a fashion similar to that of C1s to lead to the formation of the C3 convertase enzyme C4b2a. MASP1 may be able to cleave C3 directly. The alternative pathway is initiated by the covalent binding of a small amount of C3b to hydroxyl groups on cell-surface carbohydrates and proteins and is activated by low-grade cleavage of C3 in plasma. This C3b binds factor B, a protein homologous to C2, to form a C3bB complex. Factor D cleaves factor B bound to C3b to form the alternative pathway C3 complex C3bBb. The binding of properdin stabilizes this enzyme. The C3 convertase enzymes cleave many molecules of C3 to C3b, which bind covalently around the site of complement activation. Some of this C3b binds to the C4b and C3b in the convertase enzymes of the classical and alternative pathways, respectively, forming C5 convertase enzymes. This C3b acts as an acceptor site for C5, which is cleaved to form the anaphylatoxin C5a and C5b, which initiates the formation of the membrane-attack complex. The activities of biologically active proteins and protein fragments of the complement pathway are described in (Table 1).")
63
Figure 2. Regulation of the Cleavage of C3 by Factor H and Factor I.
The first product of the cleavage of C3 by a C3 convertase is C3b, which has an activated internal thioester bond. This bond enables C3b to bind covalently to hydroxyl groups on nearby carbohydrates and protein-acceptor groups. If the acceptor molecule is on a host cell surface, then protective regulatory mechanisms come into play. This is illustrated by the binding of factor H to C3b, which acts as a cofactor to the serine esterase factor I. Factor I cleaves the C3 into an inactive product, iC3b, releasing a small peptide, C3f. The iC3b can no longer participate in the formation of a C3 convertase enzyme. If C3b binds covalently to a bacterium, then the enzyme precursor factor B binds to the C3b. Factor B that is bound to C3b is susceptible to cleavage and activation by the enzyme factor D. This leads to the formation of the C3 convertase enzyme C3bBb, which is stabilized by the binding of properdin. This enzyme cleaves more C3, leading to the deposition of additional C3b on the bacterium. The carbohydrate environment of the surface on which the C3b is deposited determines the relative affinity of C3b for factor H or factor B. On host cell surfaces bearing polyanions such as sialic acid, factor H binds to C3b with a higher affinity than does factor B. On microbial surfaces that lack a polyanionic coating, factor B binds to C3b with a higher affinity than does factor H, leading to amplified cleavage of C3.

64
Figure 3. Glomerulonephritis in the Presence of C3 Nephritic Factor.
Panel A shows a glomerulus from a patient with type II membranoproliferative glomerulonephritis (hematoxylin and eosin, x200). There is mesangial expansion and hypercellularity, with thickening of glomerular capillary walls. Panel B shows the characteristic electron-dense deposits in the glomerular basement membrane (arrows) (x12,000). Photographs courtesy of Dr. Terry Cook, Department of Histopathology, Imperial College School of Medicine, Hammersmith Hospital, London
. There is mesangial expansion and hypercellularity, with thickening of glomerular capillary walls. Panel B shows the characteristic electron-dense deposits in the glomerular basement membrane (arrows) (x12,000). Photographs courtesy of Dr. Terry Cook, Department of Histopathology, Imperial College School of Medicine, Hammersmith Hospital, London.")
65
Figure 4. Facial Rash in a Patient with a Hereditary C3 Deficiency.
This rash occurred each time the patient had a pyogenic infection of the respiratory tract, and in each instance, the rash lasted a few days.

66
Figure 5. The Waste-Disposal Hypothesis for Systemic Lupus Erythematosus.
In Panel A, a macrophage is shown engulfing an apoptotic cell. There are a variety of ligands on apoptotic cells and receptors on macrophages that make this process extremely efficient. The binding of C1q, C-reactive protein, and IgM to apoptotic cells may promote the activation of complement, leading to the clearance of apoptotic cells by ligation of complement receptors. The binding of serum amyloid P component masks autoantigen on the surface of apoptotic cells and promotes their safe disposal. Once the macrophage has engulfed the apoptotic cell, it secretes the antiinflammatory cytokine transforming growth factor (TGF- ). As shown in Panel B, when there is an excess of apoptotic cells and the failure of one or more of the normal systems of receptor–ligand recognition for the uptake of apoptotic cells, immature dendritic cells may take up apoptotic cells. If this occurs in the presence of inflammatory cytokines such as granulocyte–macrophage colony-stimulating factor (GM-CSF), tumor necrosis factor (TNF- ), and interleukin-1, the dendritic cell may mature into an autoantigen-presenting cell. The dendritic cell is shown presenting autoantigens to a T cell in the presence of costimulatory molecules and cytokines. Panel C shows an autoreactive B cell that has taken up autoantigens from an apoptotic cell through its antibody receptors. The B cell is receiving help from an activated T cell, which is expressing costimulatory molecules and cytokines involved in the maturation of B cells, including an important member of the tumor necrosis family, B lymphocyte stimulator (BLyS), also referred to as zTNF-4. The autoreactive B cell divides and matures into a plasma cell that secretes autoantibodies. It is likely that in the majority of patients, systemic lupus erythematosus develops only in the presence of abnormalities in more than one of these steps.
. As shown in Panel B, when there is an excess of apoptotic cells and the failure of one or more of the normal systems of receptor–ligand recognition for the uptake of apoptotic cells, immature dendritic cells may take up apoptotic cells. If this occurs in the presence of inflammatory cytokines such as granulocyte–macrophage colony-stimulating factor (GM-CSF), tumor necrosis factor (TNF- ), and interleukin-1, the dendritic cell may mature into an autoantigen-presenting cell. The dendritic cell is shown presenting autoantigens to a T cell in the presence of costimulatory molecules and cytokines. Panel C shows an autoreactive B cell that has taken up autoantigens from an apoptotic cell through its antibody receptors. The B cell is receiving help from an activated T cell, which is expressing costimulatory molecules and cytokines involved in the maturation of B cells, including an important member of the tumor necrosis family, B lymphocyte stimulator (BLyS), also referred to as zTNF-4. The autoreactive B cell divides and matures into a plasma cell that secretes autoantibodies. It is likely that in the majority of patients, systemic lupus erythematosus develops only in the presence of abnormalities in more than one of these steps.")
67
Tolerance and Autoimmune

68
Figure 1. Central Mechanisms of the Induction of Tolerance
Figure 1. Central Mechanisms of the Induction of Tolerance. Immature T cells migrate to the thymus, where they encounter antigen presented by thymic epithelial cells. Cells whose T-cell receptors have a low affinity for the complex of self peptide and a self major-histocompatibility-complex (MHC) molecule do not receive a signal to switch off the process of spontaneous apoptosis and therefore die in the thymus. Cells whose T-cell receptors have a high affinity for such complexes are also eliminated by means of apoptosis. The remaining T cells have an intermediate affinity for these complexes, and these mature in the thymus and migrate to the periphery, where they can become activated.
 molecule do not receive a signal to switch off the process of spontaneous apoptosis and therefore die in the thymus. Cells whose T-cell receptors have a high affinity for such complexes are also eliminated by means of apoptosis. The remaining T cells have an intermediate affinity for these complexes, and these mature in the thymus and migrate to the periphery, where they can become activated.")
69
Figure 2. Peripheral Mechanisms of the Induction of Tolerance
Figure 2. Peripheral Mechanisms of the Induction of Tolerance. T cells that are physically separated from their specific antigen — for example, by the blood–brain barrier — cannot become activated, a circumstance referred to as immunologic ignorance. T cells that express the Fas (CD95) molecule on their surface can receive their signals from cells that express Fas ligand and undergo apoptosis, a process known as deletion. One example of inhibition is as follows: CD152 binds CD80 on antigen-presenting cells, thereby inhibiting the activation of T cells. Regulatory T cells can inhibit, or suppress, other T cells, most likely through the production of inhibitory cytokines such as interleukin-10 and transforming growth factor (TGF- ).
 molecule on their surface can receive their signals from cells that express Fas ligand and undergo apoptosis, a process known as deletion. One example of inhibition is as follows: CD152 binds CD80 on antigen-presenting cells, thereby inhibiting the activation of T cells. Regulatory T cells can inhibit, or suppress, other T cells, most likely through the production of inhibitory cytokines such as interleukin-10 and transforming growth factor (TGF- ).")
Presentasi serupa

































>")
 : ADALAH CABANG MATEMATIKA YANG MEMPELAJARI PENGATURAN OBJEK- OBJEK. ADALAH CABANG.>")
 (Part 2)>")
