Download presentation
1
Katabolisme Protein dan Lipid
Kelompok 5 Megawati Wahyu S. ( ) Ria Sherly Farida ( ) Fenly Arismaya ( )
 Ria Sherly Farida ( ) Fenly Arismaya ( )")
2
KATABOLISME PROTEIN

3
FUNGSI PROTEIN ANTARA LAIN:
sebagai enzim sebagai hormon sebagai alat transpor dalam darah, misalnya transferrin : transpor zat besi ikut mempertahankan keseimbangan asam basa cairan tubuh sebagai antibodi

4
Protein dibedakan menjadi:
Protein struktural adalah protein yang Berfungsi struktural misalnya kolagen, Keratin. Protein fungsional adalah yg mengerjakan Fungsi tertentu misalnya enzim, hormon.

5
Metabolisme Protein Meliputi: Degradasi protein (makanan dan protein intraseluler) menjadi asam amino Oksidasi asam amino Biosintesis asam amino Biosintesis protein

6
I. Degradasi Protein menjadi Asam amino
Protein dalam makanan dapat berasal dari hewan dan tumbuhan yang dikonsumsi manusia. Pemecahan protein dalam mulut Tidak ada enzim dalam mulut untuk mendregadasi protein. Pemecahan protein dalam perut (lambung) HCL disekresi oleh lendir-lendir mukosa mendestabilkan struktur
 HCL disekresi oleh lendir-lendir mukosa mendestabilkan struktur.")
7
Protein yang telah bereaksi dengan pepsin menghasilkan oligopeptida.
Pemecahan dalam usus 12 jari oligopeptida kemudian akan bereaksi dengan beberapa enzim seperti enzim tripsin, kimotripsin elastase membentuk senyawa peptida

8
Stadium I, protein, karbohidrat dan polisakarida dipecah menjadi molekul pembangun Stadium II, seluruh molekul pembangun terdegradasi menjadi hasil yang umum, yaitu kelompok asetil dari asetil Ko-A Hasil umum dari proses degradasi / penguraian Stadium III, katabolisme bergabung melalui siklus asam sitrat (TCA) menghasilkan H2O, CO2 dan amonia Hasil akhir molekul sederhana
 menghasilkan H2O, CO2 dan amonia Hasil akhir molekul sederhana")
9
Metabolisme Asam Amino

10
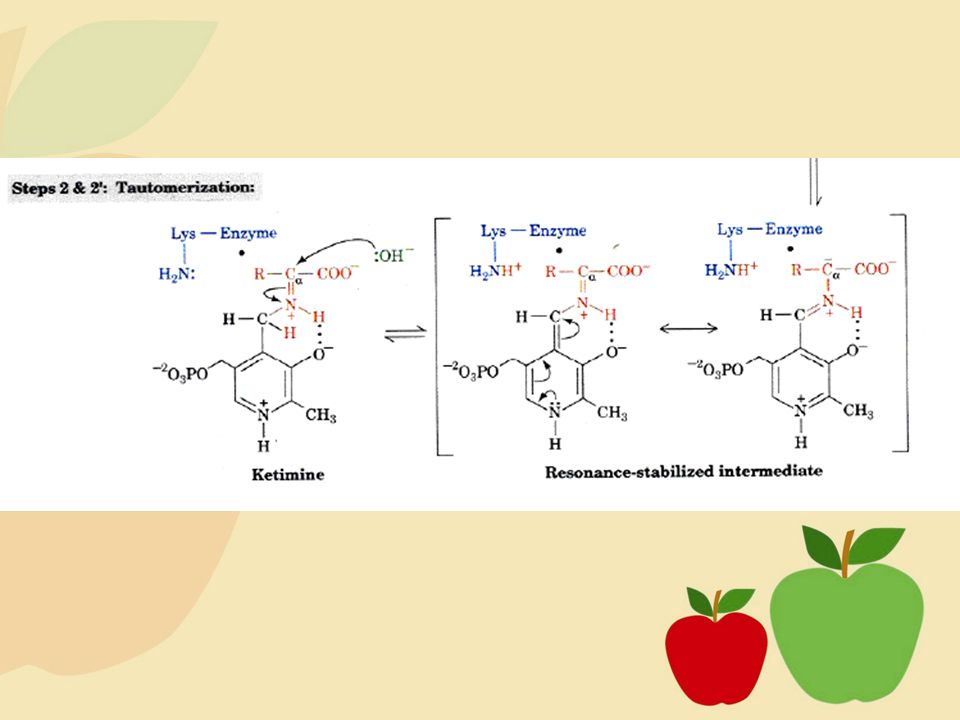
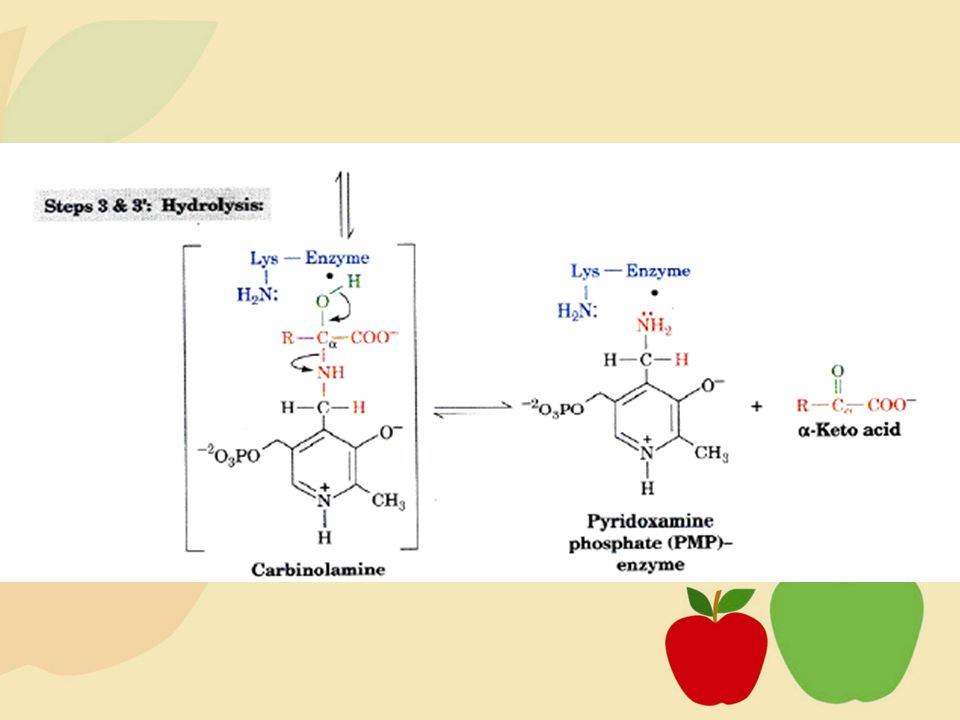
Tahap awal metabolisme asam amino melibatkan pelepasan gugus amino, kemudian baru perubahan kerangka karbon pada molekul asam amino. Dua proses utama pelepasan gugus amino, yaitu: transaminasi, yaitu proses katabolisme asam amino yang melibatkan gugus amino dari satu asam amino kepada asam amino lain, deaminasi oksidatif yang menggunakan enzim dehidrogenase sebagai katalis (Poedjiadi, 1994: ).
.")
11
Transaminasi

12
Mekanisme Reaksi

15
Deaminasi Oksidatif

16
Reaksi Keseluruhan

17
Siklus Urea

18
Reaksi-reaksi dalam Siklus Urea
Carbamoyl phosphate synthetase (Regulating enzyme)
")
20
2. Ornithine transcarbamoylase
Merupakan reaksi transfer gugus carbomyl (O=C-NH2) menjadi ornithine untuk memproduksi citrulline 3. Argininosuccinate Synthetase
 menjadi ornithine untuk memproduksi citrulline. 3. Argininosuccinate Synthetase.")
21
4. ARGININOSUCCINASE 5. ARGINASE
Merupakan eliminasi arginin dari aspartat menjadi fumarat 5. ARGINASE Hidrolisis arginin membentuk urea dan meregenerasi ornithin

22
Keseluruhan Reaksi Siklus Urea

23
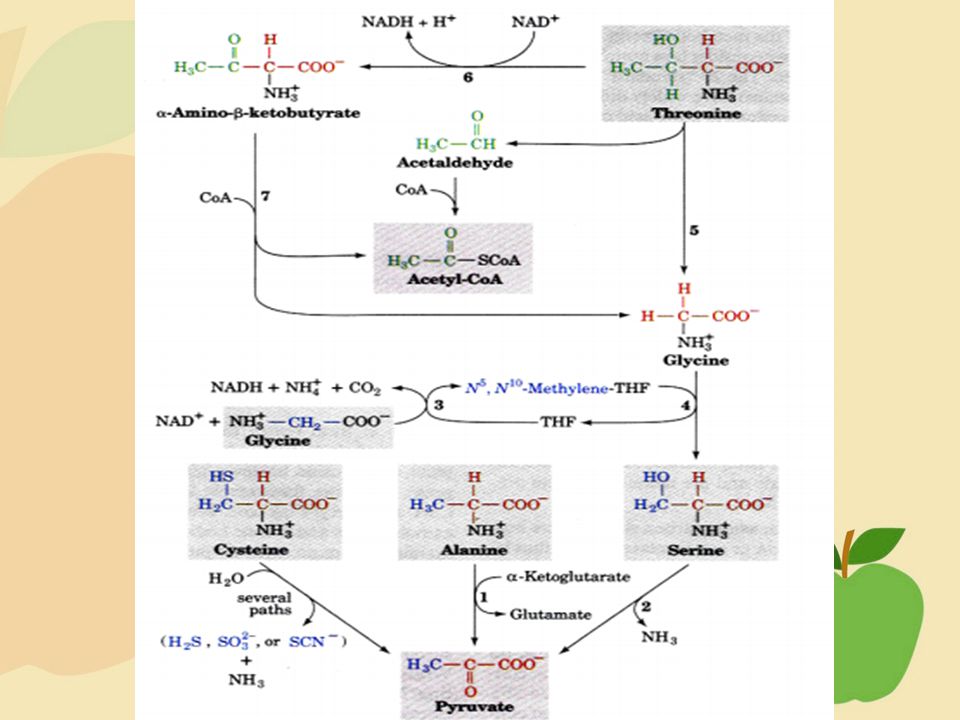
REAKSI ASAM AMINO YANG AKAN MASUK
DALAM SIKLUS KREB

25
Tahap Pembentukan Asetil KoA dari piruvat
Pada proses pembentukan asetil koa ini, terjadi subtitusi gugus Ester piruvat menjadi gugus (S-CoA). Proses ini juga disertai pembentukan NAD mnjadi NADH Kofaktor : senyawa bukan protein Koenzim adalah kofaktor yang terikat pada enzim longgar.
. Proses ini juga disertai pembentukan NAD mnjadi NADH. Kofaktor : senyawa bukan protein. Koenzim adalah kofaktor yang terikat pada enzim longgar.")
26
Piruvat + KoA + NAD+ asetil KoA + CO2 + NADH
Dekarboksilasi oksidatif piruvat menjadi asetil KoA dikatalisis oleh Kompleks Piruvat Dehidrogenase(PDC). Reaksi bersih yang terjadi adalah seperti berikut : Piruvat + KoA + NAD+ asetil KoA + CO2 + NADH Dekarboksilasi adalah reaksi kimia yang melibatkan penghapusan kelompok karboksil (-COOH) dari senyawa. Tiamin pirofosfat (TPP), lipoamida, dan FAD bertindak sebagai kofaktor katalitik, sedangkan KoA dan NAD+ merupakan kofaktor stoikiometrik.
. Reaksi bersih yang terjadi adalah seperti berikut : Piruvat + KoA + NAD+ asetil KoA + CO2 + NADH. Dekarboksilasi adalah reaksi kimia yang melibatkan penghapusan kelompok karboksil (-COOH) dari senyawa. Tiamin pirofosfat (TPP), lipoamida, dan FAD bertindak sebagai kofaktor katalitik, sedangkan KoA dan NAD+ merupakan kofaktor stoikiometrik.")
27
Mekanisme Singkat Pembentukkan Asetil KoA dari piruvat

28
Tahapan Mekanisme Pembentukkan Asetil KoA dari piruvat
1.Enzim Piruvat Dehidrogenase (E1) mengkatalisis dekarboksilasi Piruvat saat bergabung dengan TPP. Reaksi ini juga dibantu oleh beberapa kofaktor seperti Tiamin pirofosfat (TPP), lipoamida, dan FAD bertindak sebagai kofaktor katalitik, sedangkan KoA dan NAD+ merupakan kofaktor stoikiometrik. Reaksi bersihnya dapat ditulis : Piruvat + TPP hidroksietil-TPP + CO2 Struktur TPP(atas) Tiamin pirofosfat (TPP), lipoamida, dan FAD bertindak sebagai kofaktor katalitik, sedangkan KoA dan NAD+ merupakan kofaktor stoikiometrik. Pada gugus TPP, atom karbon antara atom Nitrogen dan Sulfur pada cincin tiazol (Gambar 9) merupakan karbon paling bersifat asam dibandingkan dengan sebagian besar gugus =CH lainnya.
 mengkatalisis dekarboksilasi Piruvat saat bergabung dengan TPP. Reaksi ini juga dibantu oleh beberapa kofaktor seperti Tiamin pirofosfat (TPP), lipoamida, dan FAD bertindak sebagai kofaktor katalitik, sedangkan KoA dan NAD+ merupakan kofaktor stoikiometrik. Reaksi bersihnya dapat ditulis : Piruvat + TPP hidroksietil-TPP + CO2. Struktur TPP(atas) Tiamin pirofosfat (TPP), lipoamida, dan FAD bertindak sebagai kofaktor katalitik, sedangkan KoA dan NAD+ merupakan kofaktor stoikiometrik. Pada gugus TPP, atom karbon antara atom Nitrogen dan Sulfur pada cincin tiazol (Gambar 9) merupakan karbon paling bersifat asam dibandingkan dengan sebagian besar gugus =CH lainnya.")
29
TPP mudah untuk terionisasi menjadi karbonion, yang kemudian dapat mudah untuk bergabung dengan gugus karbonil dari Piruvat. Karbon C-2 pada TPP akan bertindak sebagai nukleofil yang akan menyerang atom karbon paling positif pada Piruvat. Dibawah ini merupakan bentuk terionisasi dari TPP : Akibat karbon yang positif ini, TPP mudah untuk terionisasi menjadi karbonion (Gambar 10) yang kemudian dapat mudah untuk bergabung dengan gugus karbonil dari Piruvat. Karbon C-2 pada TPP akan bertindak sebagai nukleofil yang akan menyerang atom karbon paling positif pada Piruvat (Gambar 11). Cncin Nitrogen TPP yang bermuatan positif kemudian akan bertindak sebagai penampung negatif untuk menstabilkan pembentukan muatan egative untuk dekarboksilasi (Gambar 12). Protonasi kemudian dapat membentuk hidroksietil tiamin pirofosfat yang dapat beresonansi.
 yang kemudian dapat mudah untuk bergabung dengan gugus karbonil dari Piruvat. Karbon C-2 pada TPP akan bertindak sebagai nukleofil yang akan menyerang atom karbon paling positif pada Piruvat (Gambar 11). Cncin Nitrogen TPP yang bermuatan positif kemudian akan bertindak sebagai penampung negatif untuk menstabilkan pembentukan muatan egative untuk dekarboksilasi (Gambar 12). Protonasi kemudian dapat membentuk hidroksietil tiamin pirofosfat yang dapat beresonansi.")
30
Mekanisme Tahap Penyerangan TPP pada Piruvat
TPP menyerang piruvat - C menyerang karbokation pada piruvat. Resonansi penghilangan CO2 Setelah penghilangan CO2, diresonasi kembali sehingga dihasilkan produk ini (hidroksi etil TPP)
")
31
3. Enzim Piruvat Dehidrogenase (E1) kembali berperan dalam mengkatalisis reaksi. Gugus hidroksietil yang terikat pada TPP teroksidasi membentuk gugus asetil yang kemudian ditransfer ke lipoamida menjadi Asetil Lipoamida. Pada reaksi akan terbentuk pula karbanion TPP. Lipoamida kofaktor katalitik Perpindahan gugus hidroksietil ke lipoamida menjadi acetil poamida

32
4. Gugus asetil ditransfer dari asetil lipoamida ke kofaktor KoA untuk membentuk Asetil KoA yang dikatalisis oleh Dihidrolipoil transasetilase (E2).
..")
33
Katabolisme Lipid

34
Lipid Lipid adalah senyawa organik yang meliputi lemak, minyak dan lilin. Tidak larut dalam air tetapi larut dalam pelarut nonpolar seperti eter, kloroform dan benzena. Lipid didistribusikan di semua organ, terutama di jaringan adiposa di mana lipid mewakili lebih dari 90 persen sitoplasma sel.

35
Pencernaan Perubahan makanan menjadi bentuk yang dapat diserap oleh tubuh disebut pencernaan / digestion. Semua enzim yang terlibat dalam pencernaan larut dalam air, sedangkan lipid tidak dapat larut dalam air. Masalah ini diatasi dengan emulsifikasi lemak, pemecahan lemak berukuran besar menjadi lebih kecil sehingga ukuran luas areanya bertambah, yang dapat memfasilitasi interaksi lemak dengan enzim lipase penghidrolisis lemak.

36
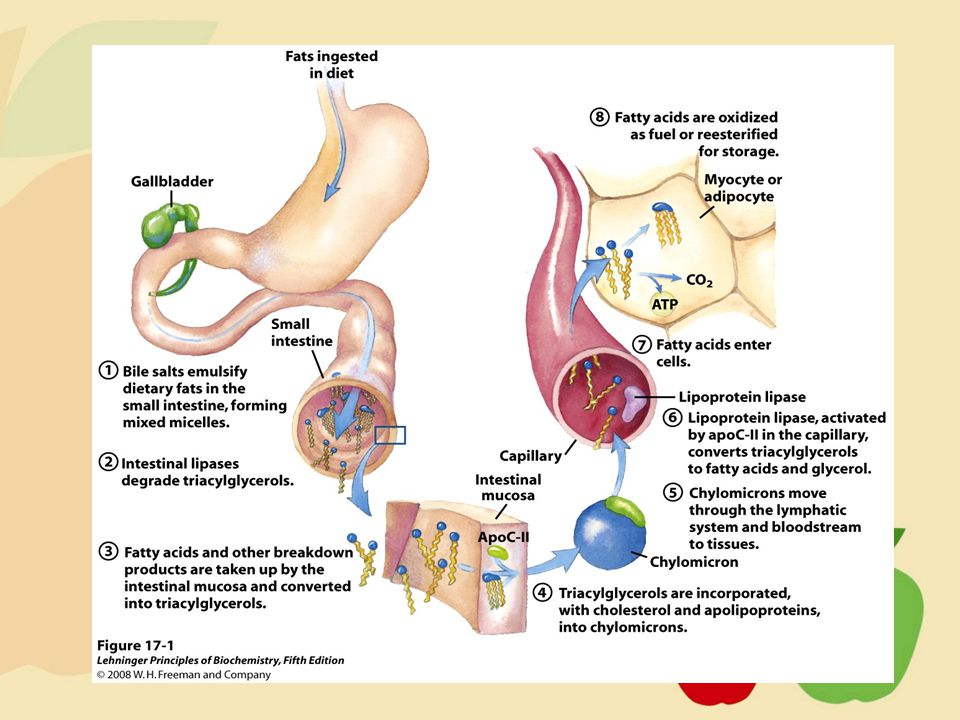
Pemecahan Lipid Emulsifikasi Lemak
Asam lemak dapat dipecah melalui hidrolisis ikatan ester trigliserida, tetapi ketidaklarutan dari trigliserida menjadi masalah. Pemecahan terjadi pada lemak dalam makanan menjadi bagian yang lebih kecil dengan luas permukaan yang cukup luas untuk bereaksi cepat dengan enzim pencernaan. Enzim lipase membantu pemecahan asam lemak pendek dalam proses yang terjadi di mulut. - Garam empedu seperti asam kolat mengandung sisi hidrofobik dan sisi hidrofilik, sehingga memungkinkan garam empedu untuk melarutkan lipid-air, dengan permukaan hidrofobik kontak dengan fase nonpolar dan permukaan hidrofilik dalam media berair. - Reaksi ini mengemulsi lemak dan menghasilkan campuran misel, yang memungkinkan serangan oleh enzim pencernaan yang larut dalam air dan memfasilitasi penyerapan lipid melalui mukosa usus. - Dengan demikian, penyerapan efisien lipid tergantung pada adanya asam empedu yang cukup untuk melarutkan lipid

37
Pemecahan Lipid 2. Pencernaan Lemak Emulsifikasi lemak membuatnya rentan terhadap hidrolisis oleh enzim yang disekresikan pankreas. Enzim paling penting yang terlibat adalah lipase pankreas. Lipase pankreas spesifik untuk hidrolisis ikatan ester primer, ikatan ester 1 atau 3, tapi bukan ikatan posisi sentral 2. Sebagai hasil dari konversi ini, 2-monogliserida (2-monoasilgliserol) adalah produk akhir utama pencernaan trigliserida. Kurang dari 10% dari trigliserida tetap tidak terhidrolisis dalam usus.
 adalah produk akhir utama pencernaan trigliserida. Kurang dari 10% dari trigliserida tetap tidak terhidrolisis dalam usus.")
39
Penyerapan Lemak Asam lemak bebas yang diserap oleh sel-sel dinding usus halus melalui difusi sederhana. Sebagai asam lemak yang masuk dengan cepat diubah menjadi trigliserida. Sebagian dari gliserol dan asam lemak rantai pendek secara langsung diserap oleh sirkulasi portal dan dibawa ke hati. Gliserol dan asam lemak yang masuk ke sel-sel epitel usus diubah menjadi trigliserida dan dalam lakteal, ditutupi dengan lapisan fosfolipid hidrofilik, kolesterol, ester kolesterol dan apoprotein apo B. Setelah dikemas menjadi bentuk terlarut yang lebih hidrofilik, masuk ke dalam sirkulasi limfatik.

40
khilomikron Dibentuk dalam sel epitel usus
Paling banyak mengandung lipida (terutama TG) Diameternya berkisar 100 – 500 nm Mengandung apolipoprotein : Apo-B; Apo-C dan Apo-A Fungsinya mengangkut TG dari usus ke jaringan lemak dan mengangkut kholesterol bebas / kholesterol ester dari usus
 Diameternya berkisar 100 – 500 nm. Mengandung apolipoprotein : Apo-B; Apo-C dan Apo-A. Fungsinya mengangkut TG dari usus ke jaringan lemak dan mengangkut kholesterol bebas / kholesterol ester dari usus.")
41
Mobilisasi Triasilgliserol dalam Jaringan Adiposa

42
Masuknya gliserol ke dalam jalur glikolisis

43
Masuknya Asam Lemak ke dalam Mitokondria melalui Asil-karnitin

44
Konversi Asam Lemak menuju Asil-CoA

45
Tahapan Oksidasi Asam Lemak

46
Oksidasi Asam Lemak Pencernaan lemak dimulai di usus halus. Lemak diemulsi oleh garam empedu dan dihidrolisis oleh lipase pankreas untuk membentuk asam lemak bebas. Asam lemak bebas ini digabungkan dengan gliserol (diproduksi oleh proses glikolitik) untuk membentuk trigliserida, kemudian digabungkan dengan protein untuk membentuk lipoprotein dan masuk ke dalam sirkulasi untuk melakukan berbagai fungsi biolologikal seperti oksidasi, penyimpanan dan pembentukan lipid baru. Dengan demikian berbagai asam lemak mungkin ada dalam bentuk bebas serta dalam bentuk teresterifikasi (trigliserida) dalam darah. Asam lemak merupakan sumber langsung untuk oksidasi lemak dalam berbagai jaringan, yaitu hati, jaringan adiposa, otot, jantung, ginjal, otak, paru-paru.
 untuk membentuk trigliserida, kemudian digabungkan dengan protein untuk membentuk lipoprotein dan masuk ke dalam sirkulasi untuk melakukan berbagai fungsi biolologikal seperti oksidasi, penyimpanan dan pembentukan lipid baru. Dengan demikian berbagai asam lemak mungkin ada dalam bentuk bebas serta dalam bentuk teresterifikasi (trigliserida) dalam darah. Asam lemak merupakan sumber langsung untuk oksidasi lemak dalam berbagai jaringan, yaitu hati, jaringan adiposa, otot, jantung, ginjal, otak, paru-paru.")
47
Asam lemak dioksidasi menjadi CO2 dan air dengan pembebasan sejumlah besar energi. Oksidasi terjadi dalam mitokondria karena semua enzim yang dibutuhkan untuk oksidasi ada dalam mitokondria. Oksidasi asam lemak ada tiga jenis, berdasarkan posisi atom karbon yang akan dioksidasi (α, β dan γ). Namun β-oksidasi asam lemak adalah yang dominan dan banyak terjadi serta menyediakan sejumlah besar energi dibandingkan α dan γ oksidasi. β-oksidasi asam lemak, tahapan awalnya adalah aktivasi asam lemak.

48
Aktivasi Asam Lemak Asam lemak adalah molekul kimia yang relatif lembam dan karenanya harus dikonversi ke perantara aktif untuk memulai reaksi β-oksidasi. Aktivasi asam lemak terjadi di sitosol dengan adanya ATP, koenzim A dan asil CoA sintetase. Asam lemak yang diaktifkan kemudian masuk ke dalam mitokondria dengan bantuan protein pembawa, karnitin dengan adanya enzim karnitin asil transferase.

49
Aktivasi Asam Lemak Oksidasi asil CoA (asam lemak) terjadi melalui beberapa tahap yang mengarah pada pembentukan asetil CoA (C2) dan asil CoA memiliki dua atom karbon yang berkurang dari asam lemak awal dimulainya siklus β-oksidasi. Asil CoA kemudian masuk ke dalam siklus oksidasi yang sama sampai semua atom karbon dilepaskan sebagai asetil CoA. Reaksi ini memerlukan kofaktor seperti Flavin Adenin Dinucleotide (FAD) dan Nicotinamide Adenine Dinucleotide (NAD +).
 terjadi melalui beberapa tahap yang mengarah pada pembentukan asetil CoA (C2) dan asil CoA memiliki dua atom karbon yang berkurang dari asam lemak awal dimulainya siklus β-oksidasi. Asil CoA kemudian masuk ke dalam siklus oksidasi yang sama sampai semua atom karbon dilepaskan sebagai asetil CoA. Reaksi ini memerlukan kofaktor seperti Flavin Adenin Dinucleotide (FAD) dan Nicotinamide Adenine Dinucleotide (NAD +).")
50
Oksidasi Asam Lemak Disebut β-oksidasi karena karbon β dari asam lemak yang dioksidasi. β-oksidasi menghasilkan asetil-KoA and asam lemak yang kehilangan 2-karbon.

51
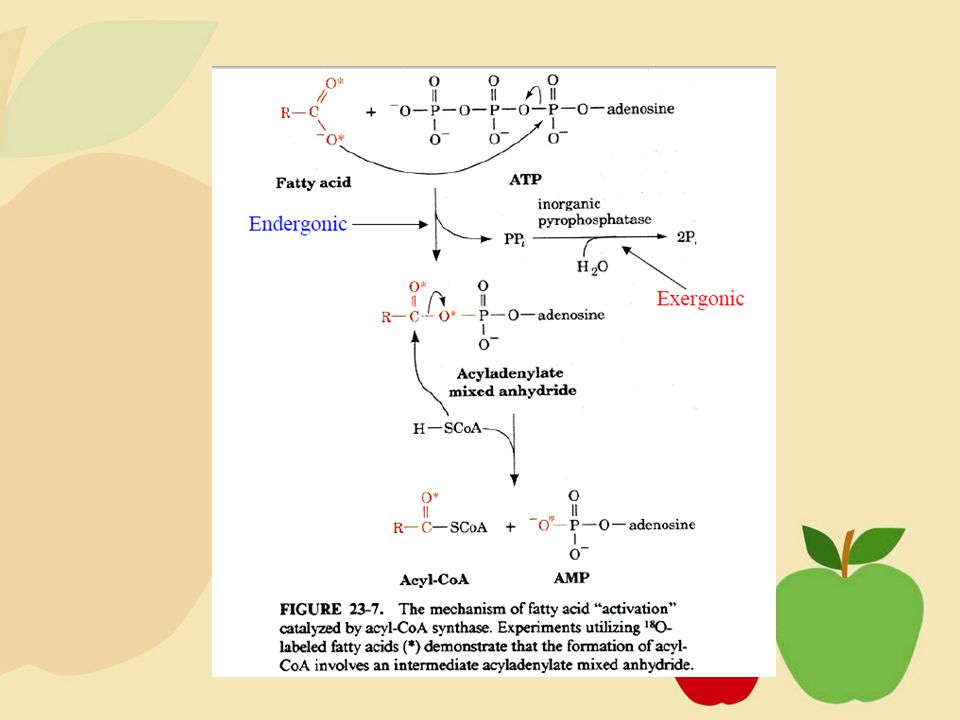
KoA menyerang/berikatan dengan asam lemak menggunakan hidrolisis energi ATP, dan asil-KoA adalah hasilnya. Asam lemak + KoA + ATP → Asil-KoA + AMP + PPi (→ 2Pi) Dalam reaksi ini,oksigen karboksil bermuatan negatif (⎯O-) dari asam lemak menyerang fosfat α dari ATP, dan pirofosfat dipecah dari ATP. Reaksi ini berlangsung endergonik. Reaksi subsekuen (hidrolisis pirofosfat) relatif reaksi exergonik(ΔG = kJ/mol).
 Dalam reaksi ini,oksigen karboksil bermuatan negatif (⎯O-) dari asam lemak menyerang fosfat α dari ATP, dan pirofosfat dipecah dari ATP. Reaksi ini berlangsung endergonik. Reaksi subsekuen (hidrolisis pirofosfat) relatif reaksi exergonik(ΔG = kJ/mol).")
53
Transport dari Asil-KoA yang dihasilkan di Sitosol
Asil-KoA dibuat di sitoplasma, tetapi β-oksdasi terjadi di mitokondria. Sehingga Asil-KoA harus ditransport ke dalam mitokondria. Asil-KoA tidak memiliki protein pembawa yang spesifik, maka digunakan sistem pembawa yang khusus. Awalnya asil-KoA bereaksi dengan karnitin, menghasilkan asil-karnitin dan KoA

54
Transport dari Asil-KoA yang dihasilkan di Sitosol
Asil-karnitin ditransport melewati membran mitokondria oleh protein pembawa karnitin ke dalam mitokondria. Asil-karnitin bereaksi dengan KoA dalam mitokondria dan menghasilkan asil-KoA dan karnitin.

55
β-oksidasi Desaturasi antara karbon α dan β menggunakan FAD menjadi reduksi FADH2, dan hasilnya adalah senyawa trans ∆2. Enzim : asil-KoA dehidrogenase. Hidrasi dengan H2O menjadi 3 karbon untuk menghasilkan 3-L-hidroksiasil-KoA. Enzim : enoil-KoA hidratase.

56
β-oksidasi 3. Oksidasi menggunakan NAD+ menjadi NADH, dan menghasilkan β-ketoasil-KoA. Enzim : 3-L-hidroksiasil-KoA dehidrogenase. 4. Pemecahan oleh KoA menghasilkan asetil-KoA dan asil-KoA dengan karbon berkurang 2. Enzim : β-ketoasil-KoA tiolase.

57
Jalur -oksidasi









 Sistem sitokhrom.>")











>")
 INDAH PURWANTI (121810301064) OCTAVIANTI NURYANI (121810301067)>")
