METABOLISME ASAM LEMAK
Sumber lemak : Makanan Biosintesis de novo Simpanan tubuh adiposit Masalah utama sifatnya tidak larut dalam air. Lemak diemulsi oleh garam empedu – disintesis oleh liver & disimpan dlm empedu mudah dicerna & diserap Transportasi membentuk kompleks dg protein lipoprotein
Garam empedu terdiri dr asam empedu yg berasal dari kolesterol Garam empedu bersifat amfifatik mengemulsi lemak membentuk misel Lemak dipecah oleh lipase pankreas hasil?
Struktur & tatanama asam lemak penamaan sistematis : nama hk induk huruf -a terakhir diganti –oat Mis. as lemak jenuh C18 (18:0) : as oktadekanoat (18:2) : ada 2 ikt rangkap Posisi ikt rangkap Mis sis-9 : ada terdapat ikt rangkap sis antara atom C 9 dan 10 trans-2 :ada ik rangkap trans antara atom C 2 dan 3. Cara lain menunjukkan posisi ikatan rangkap : dengan menghitung dari ujung distal dengan atom karbon (karbon metil) diberi nomor 1 Mis Asam lemak -3
Beberapa Jenis Asam Lemak Jumlah Karbon Ikatan rangkap Nama umum Nama sistematik Rumus 12 14 16 18 20 22 24 Laurat Miristat Palmitat Stearat Arakidat Behenat Lignoserat n-Dodekanoat n-Tetradekanoat n-Heksadekanoat n-Oktadekanoat n-Eikosanoat n-Dokosanoat n-Tetrakosanoat CH3(CH2)10COO- CH3(CH2)12COO- CH3(CH2)14COO- CH3(CH2)16COO- CH3(CH2)18COO- CH3(CH2)20COO- CH3(CH2)22COO- 1 2 3 4 Palmitoleat Oleat Linoleat Linolenat Arakidonat sis-9-heksadesenoat sis-9-oktadesenoat sis- sis-9-12-oktadekadienoat semua sis-9-12-15-oktadekatrionat semua sis-5-8-11-14-eikosatetraenoat CH3(CH2)5CH=CH(CH2)7COO- CH3(CH2)7CH=CH(CH2)7COO- CH3(CH2)4(CH=CHCH2)2(CH2)6COO- CH3CH2(CH=CHCH2)3(CH2)6COO- CH3(CH2)4(CH=CHCH2)4(CH2)2COO-
Triasilgliserol bentuk simpanan energi metabolisme yang pekat berada dalam bentuk tereduksi dan anhidrat Perolehan energi : oksidasi sempurna asam lemak : 9 kcal g-1 (38 kJ g-1) karbohidrat dan protein hanya : 4 kcal g-1 (17 kJ g-1) Pada sel mamalia, tempat akumulasi triasilgliserol adalah sitoplasma dari sel-sel adiposa (sel lemak). Tetesan-tetesan atau butiran-butiran triasilgliserol bergabung membentuk gumpalan besar yang dapat menempati sebagian besar volume sel lemak
PROSES PEMAKAIAN ASAM LEMAK SBG BAHAN BAKAR 3 tahap : Mobilisasi triasilgliserol Aktivasi dan transportasi asam lemak Pemecahan asam lemak menjadi asetil koA (β-oksidasi)
Mobilisasi asam lemak Hidrolisis triasilgliserol menjadi asam lemak dan gliserol di dalam sel lemak pelepasan asam lemak dari sel lemak, ditransport ke jaringan-jaringan yang memerlukan energi
Hidrolisis triasilgliserol
Aktivasi enzim lipase Enzim lipase dalam jaringan adiposa (jaringan lemak) diaktivasi oleh hormon-hormon : epinefrin, norepienfrin, glukagon, dan adrenokortikotropik. Hormon-hormon tsb merangsang reseptor 7TM yang mengaktivasi adenilat siklase sehingga cAMP meningkat, yang akan mengaktifkan protein kinase A, selanjutnya mengaktifkan lipase dengan cara fosforilasi
Metabolisme gliserol Gliserol yang terbentuk pada lipolisis diabsorpsi oleh liver difosforilasi dan dioksidasi menjadi dihidroksiaseton fosfat diisomerisasi menjadi gliseraldehid-3-fosfat Jadi gliserol dapat diubah menjadi piruvat atau glukosa di hati.
Metabolisme asam lemak β-oksidasi asam lemak Tahapan : Aktivasi asam lemak Transport asil lemak koA (Fatty Acyl CoA) Reaksi-reaksi : Oksidasi Hidrasi Pemutusan ikatan C-C (reaksi thiolisis)
Aktivasi Asam Lemak Asam lemak dioksidasi di mitokondria Asam lemak mengalami aktivasi sebelum memasuki mitokondria ATP memacu pembentukan ikatan tioester antara gugus karboksil asam lemak dan gugus sulfhidril pada KoA Reaksi aktivasi berlangsung di membran luar mitokondria dikatalis oleh enzim asil KoA sintetase
Reaksi: FA + CoA + ATP asil lemak koA + AMP + 2Pi + 34 kJ/mol
Trasportasi asil-koA Gugus asil pada asil-koA ditransfer ke gugus OH karnitin membentuk asil karnitin yg dikatalis karnitin asiltransferase I pd membran luar mitokondria Asil karnitin melintasi membran dalam mitokondria yg dikatalis enzim translokase Gugus asil ditransfer kembali ke koA yg berada dalam matriks mit. yg dikatalis karnitin asiltransferase II enzim translokase memindah kembali karnitin ke sitosol
Trasportasi ester Asil-koA Rate-limiting step of FA oxidation
Reaksi β oksidasi Terdiri dari 4 proses utama: Dehidrogenasi Hidratasi Thiolisis Berapakah jumlah reaksi yang dibutuhkan untuk mengoksidasi asam palmitat menjadi asetil Co A?
Step 1 : dehidrogenasi / oksidasi Berperan pada pembentukan rantai ganda antara atom C2 – C3. Mempunyai akseptor hidrogen FAD+. Antara asam lemak yg berbeda panjangnya beda enzimnya,
Step2 : Hidrasi Mengkatalisis hidrasi trans enoyl CoA Penambahan gugus hidroksi pada C no. 3 Ensim bersifat stereospesifik Menghasilkan 3-L-hidroksiasil Co. A
Step 3 : dehidrogenasi/ oksidasi Mengkatalisis oksidasi -OH pada C no. 3 / C β menjadi keton Akseptor elektronnya : NAD+
Step 4 : thiolisis β-Ketothiolase mengkatalisis pemecahan ikatan thioester. Asetil-koA dilepas dan tersisa asil lemak ko A yang terhubung dgn thio sistein mll ikatan tioester. Tiol HSCoA menggantikan cistein thiol, menghasilkan asil lemak-koA (dengan pemendekan 2 C)
β-oksidasi asam palmitat
(a) Repeat Sequence
Perolehan ATP pada oksidasi asam lemak Energi yang diperoleh pada oksidasi asam lemak dapat dihitung berdasarkan stoikhiometri setiap siklus sebagai berikut: asilKoA dipendekkan sebanyak 2 karbon dengan pelepasan FADH2, NADH dan asetil KoA Reaksi : Cn-asil KoA + FAD + NAD+ + H2O + KoA Cn-2-asil KoA + FADH2 + NADH + asetil KoA + H+
Perolehan ATP pada Oksidasi Asam Palmitat : Pemecahan palmitoil KoA (C18-asil KoA) : perlu 7 daur reaksi Pada daur ke -7, C4-ketoasil KoA mengalami tiolisis menjadi dua molekul asetil KoA Palmitoil KoA + 7 FAD + 7 NAD+ + 7 KoA + 7 H2O → 8 asetil KoA + 7 FADH2 +7 NADH + 7 H+ Pembentukan ATP : Oksidasi NADH → 2,5 ATP FADH2→ 1,5 ATP asetil KoA → 10 ATP Jumlah ATP yang terbentuk pada oksidasi palmitoil KoA : 108 10,5 dari 7 FADH2 17,5 dari 7 NADH 80 dari 8 mol asetil KoA Dua ikatan fosfat energi tinggi dipakai untuk mengaktifkan palmitat (ATP → AMP + 2 Pi) Jadi oksidasi sempurna satu mol palmitat menghasilkan 106 ATP
Degradasi asam lemak tak jenuh Membutuhkan 2 enzim tambahan yi Enoyl CoA isomerase 2,4 dienoyl CoA reduktase
Degradasi asam lemak dengan jumlah atom C ganjil Degradasi FA dgn jumlah C ganjil pd akhir beta oksidasi asetoasetil Ko A dipecah akan menghasilkan propionil Ko A dan Asetil Ko A Propionil Ko A diubah menjadi metilmalonil Ko A suksinil KoA TCA
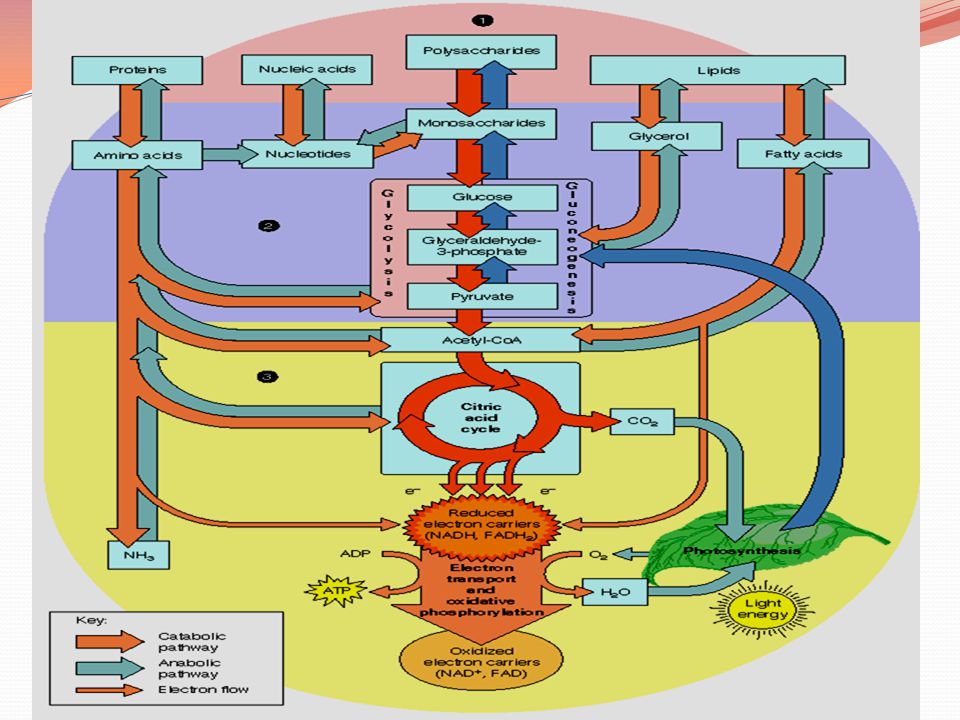
Penggunaan Asetil KoA Asetil KoA yang terbentuk pada oksidasi asam lemak dapat masuk ke dalam siklus asam sitrat hanya apabila degradasi lemak dan degradasi karbohidrat berjalan seimbang. Proses masuknya asetil KoA ke dalam siklus asam sitrat tergantung pada keberadaan oksaloasetat dari sitrat. Konsentrasi oksaloasetat rendah apabila karbohidrat tidak tersedia atau digunakan secara berlebihan. Secara normal aksaloasetat dihasilkan dari piruvat (produk glikolisis) oleh enzim piruvat karboksilase.
Pembentukan badan keton Selama puasa atau pada diabetes oksaloasetat dikonsumsi untuk menghasilkan glukosa melalui jalur glukoneogenesis, sehingga tidak ada yang dapat digunakan untuk kondensasi dengan asetil KoA. asetil KoA diubah menjadi asetoasetat dan D-3-hidroksibutirat. Senyawa-senyawa asetoasetat, D-3-hidroksibutirat dan aseton dinamakan badan-badan keton. Penderita diabetes yang tidak diobati, maka badan-badan keton ditemukan dalam darahnya dengan kadar yang tinggi.
Badan-badan keton Tempat pembentukan asetoasetat dan D-3-hidroksibutirat : liver Senyawa ini berdifusi dari mitokondria liver menuju darah kemudian ditransport ke jaringan-jaringan perifer. Otot jantung dan korteks ginjal menggunakan asetoasetat sebagai pengganti glukosa Otak juga dapat beradaptasi ketika dalam kondisi berpuasa atau diabetes sehingga dapat menggunakan asetoasetat Selama puasa jangka lama, 75 % bahan bakar yang diperlukan otak dipenuhi oleh badan-badan keton.
Reaksi pembentukan badan keton
Reaksi degradasi badan keton 3-hidroksibutirat dioksidasi menghasilkan asetoasetat dan NADH (selanjutnya diproses di rantai fosforilasi oksidatif menghasilkan energi) Asetoasetat diaktivasi melalui transfer KoA dari suksinil KoA membentuk asetoasetil KoA oleh enzim KoA transferase. Kemudian asetoasetil KoA didegradasi oleh tiolase menghasilkan asetil KoA (siap diproses di siklus asam sitrat untuk menghasilkan energi)
Sintesis Asam Lemak Tidak sepenuhnya merupakan kebalikan dari degradasi asam lemak Enzim yang berbeda bekerja dlm reaksi yang berlawanan : degradasi vs biosintesis
Perbedaan jalur sintesis dan degradasi asam lemak Sintesis asam lemak Degradasi asam lemak Lokasi Terjadi di sitosol Terjadi di matriks mitokondria Bentuk senyawa antara Terikat secara kovalen pada karier gugus asil yang dinamakan ACP (acyl carier protein) Terikat secara kovalen pada Koenzim A (KoA) Enzim-enzim yang terlibat Berasosiasi dalam sebuah rantai polipeptida yang dinamakan fatty acid synthase Tidak berasosiasi Kebutuhan oksidator / reduktor Memerlukan senyawa reduktor NADPH Memerlukan senyawa oksidator NAD+ dan FAD
Sintesis Asam Lemak Sintesis Asam lemak pada eukariotik dan prokariotik : sama Biosintesis terdiri dari 3 langkah : Biosintesis asam lemak dari asetil CoA (di sitosol) Pemanjangan rantai asam lemak (di mitokondria & ER) Desaturasi (di ER) Biosintesis as lemak membutuhkan malonil Co A sebagai substrat Diperlukan ATP Reaksi biosintesis asam palmitat: Dari 8 acetyl-CoA diperlukan 7 ATP +14 NADPH Enzim untuk sintesis asam lemak : komplek fatty acid synthase
Tahapan Sintesis Asam Lemak Reaksi awal - Karboksilasi gugus asetil menjadi malonil-KoA - Reaksi dikatalis oleh asetil KoA karboksilase Biotin-enzim + ATP + HCO3- CO2-biotin-enzim + ADP + Pi CO2-biotin-enzim + asetil KoA malonil KoA + biotin-enzim
2. Pemanjangan rantai putaran 1: pembentukan asetil ACP dan malonil ACP reaksi dikatalis oleh asetil transasilase dan malonil transasilase Asetil KoA + ACP asetil ACP + KoA Malonil KoA + ACP malonil ACP + KoA Reaksi kondensasi Asetil ACP + malonil ACP asetoasetil ACP + ACP + CO2
Reaksi Kondensasi
Reduksi gugus keto pada C-3 menjadi gugus metilen (1) asetoasetil ACP direduksi menjadi 3-hidroksi butiril ACP. Reaksi ini memerlukan NADPH sebagai pereduksi. (2) Dehidrasi 3-hidroksi butiril ACP menjadi krrotonil ACP (merupakan trans-2 enoyl ACP). (3) Reduksi krotonil ACP menjadi butiril ACP dengan menggunakan senyawa peredusi NADPH, yang dikatalis oleh enzim enoyl ACP reduktase.
Reaksi reduksi I
Reaksi dehidrasi
Reaksi reduksi II
3. Pemanjangan rantai 2 Reaksi pemanjangan rantai putaran 2 : kondensasi buritil ACP dengan malonil ACP membentuk C6--ketoasil ACP Reaksi ini sama dengan reaksi pemanjangan rantai putaran 1. Selanjutnya pemanjangan rantai diteruskan sampai terbentuk C16 asil ACP
Stoikiometri Sintesis Asam palmitat Asetil KoA + 7 Malonil KoA + 14 NADPH + 20 H+ → palmitat + 7 CO2 + 14 NADP+ + 8 KoA + 6 H2O Reaksi tersebut memerlukan malonil KoA yang disintesis dari : 7 Asetil KoA + 7 CO2 + 7 ATP → 7 malonil KoA + 7 ADP + 7 Pi + 14 H+ Jadi stoikhiometri keseluruhan sintesis palmitat adalah: 8 Asetil KoA + 7 ATP + 14 NADPH + 6 H+ → palmitat + 14 NADP+ + 8 KoA + 6 H2O + 7 ADP + 7 Pi