Upload presentasi
Presentasi sedang didownload. Silahkan tunggu

1
ENZYME MECHANISM

2
specific activity is the amount of product formed by an enzyme in a given amount of time under given conditions per milligram of enzyme. The rate of a reaction is the concentration of substrate disappearing (or product produced) per unit time (mol L − 1s − 1) The enzyme activity is the moles converted per unit time (rate × reaction volume). Enzyme activity is a measure of quantity of enzyme present. The SI unit is the katal, 1 katal = 1 mol s-1, but this is an excessively large unit. A more practical value is 1 enzyme unit (EU) = 1 μmol min-1 (μ = micro, x 10-6). The specific activity is the moles converted per unit time per unit mass of enzyme (enzyme activity / actual mass of enzyme present). The SI units are katal kg-1, but more practical units are μmol mg-1 min-1. Specific activity is a measure of enzyme efficiency, usually constant for a pure enzyme. If the specific activity of 100% pure enzyme is known, then an impure sample will have a lower specific activity, allowing purity to be calculated. The % purity is 100% × (specific activity of enzyme sample / specific activity of pure enzyme). The impure sample has lower specific activity because some of the mass is not actually enzyme.
 per unit time (mol L − 1s − 1) The enzyme activity is the moles converted per unit time (rate × reaction volume). Enzyme activity is a measure of quantity of enzyme present. The SI unit is the katal, 1 katal = 1 mol s-1, but this is an excessively large unit. A more practical value is 1 enzyme unit (EU) = 1 μmol min-1 (μ = micro, x 10-6). The specific activity is the moles converted per unit time per unit mass of enzyme (enzyme activity / actual mass of enzyme present). The SI units are katal kg-1, but more practical units are μmol mg-1 min-1. Specific activity is a measure of enzyme efficiency, usually constant for a pure enzyme. If the specific activity of 100% pure enzyme is known, then an impure sample will have a lower specific activity, allowing purity to be calculated. The % purity is 100% × (specific activity of enzyme sample / specific activity of pure enzyme). The impure sample has lower specific activity because some of the mass is not actually enzyme.")
3
Enzyme Action Each enzyme has a unique three-dimensional shape that binds and recognizes a group of reacting molecules called substrates. The active site of the enzyme is a small pocket to which the substrate directly binds. Some enzymes are specific only to one substrate; others can bind more than one substrate.

4
Enzyme-Substrate Binding

5
Models of Enzyme Action
Early theory: lock-and-key model. Active site (lock) had the same shape as the substrate (key). Only the right shape key could bind. Current theory: induced fit model. Active site closely resembles but does not exactly bind the substrate. Allows for more flexibility in type of substrate Also explains how the reaction itself occurs. As the substrate flexes to fit the active site, bonds in the substrate are flexed and stressed -- this causes changes/conversion to product.
 had the same shape as the substrate (key). Only the right shape key could bind. Current theory: induced fit model. Active site closely resembles but does not exactly bind the substrate. Allows for more flexibility in type of substrate. Also explains how the reaction itself occurs. As the substrate flexes to fit the active site, bonds in the substrate are flexed and stressed -- this causes changes/conversion to product.")
6
Molecular Recognition
How does an enzyme bind a substrate, reduce the activation barrier, and produce a product? Lock & Key Hypothesis Induced Fit Hypothesis vs.

7
C. Factors Affecting Enzyme Activity
Enzyme activity is defined as how fast an enzyme catalyzes its reaction. Many factors affect enzyme activity: Temperature: most have an optimum temp around 37oC pH: most cellular enzymes are optimal around physiological pH, but enzymes in the stomach have a lower optimum pH Concentration of enzyme and substrate: have all of the enzyme molecules been used up, even though substrate is still available?

8
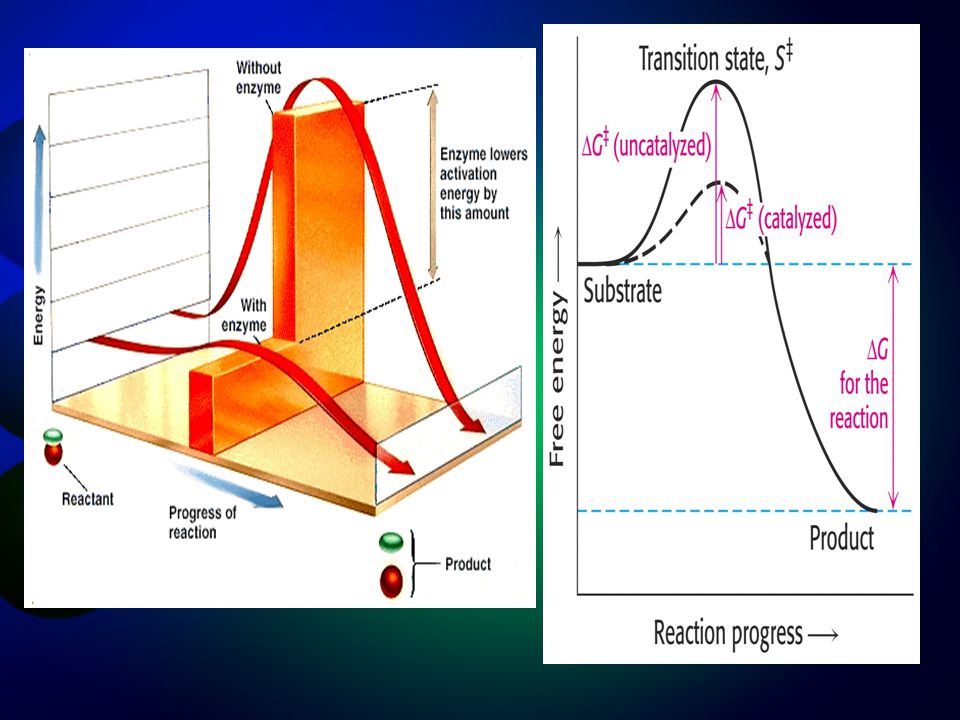
Energy of activation: ΔG‡
Effect of catalysis ΔG‡ ΔGcat‡ A → B Effect of temp ΔGT1‡ ΔGT2‡ (T1 > T2) A → B
 A → B.")
10
Rate acceleration: mechanisms
E + S ES EX‡ E + P Stabilization of the transition state: covalent bonds, metals, acid-base, and proximity. Destabilization of ES: strain, charge, electrostatics Reduced entropy in ES formation.

11
Rate acceleration: mechanisms
hydrolysis of a β-glycosidic bond yielding a unit of α-glucose

12
Major factors: pH, ions, & temp
At pH ~ 7 amino acids exist as zwitterions. The R group determines pH. aspartic acid [pKa = 4.0] arginine [pKa = 12.5]

13
Major factors: pH, ions, & temp
ionic strength temperature barley α-amylase activity plotted as a function of pH

14
Major factors: pH, ions, & temp
ionic strength temperature Having the correct ions is important. Why? barley α-amylase isozyme 1 [crystallized with Ca2+ (green)]
]")
15
Major factors: pH, ions, & temp
ionic strength temperature barley α-amylase with CaCl2 barley α-amylase w/o CaCl2

16
Michaelis-Menten Kinetics
E + S ↔ ES → E + P k1 k-1 k2 Assumptions: [1] Steady-state of the intermediate complex ES [2] Neglect back rxn from product (k-2; not shown) [3] Conservation of mass ([ET] = [E] + [ES]) Vmax = k2[ET] ν = Vmax [S] Km + [S] where: Km = (k-1 + k2) k1
 [3] Conservation of mass ([ET] = [E] + [ES]) Vmax = k2[ET] ν = Vmax [S] Km + [S] where: Km = (k-1 + k2) k1.")
17
Michaelis-Menten Kinetics

18
Michaelis-Menten Kinetics
Many types of inhibition can be included in the MM model as well as multiple substrates and steps: Inhibition: competitive (rev) noncompetitive (rev) mixed (rev) irreversible Reaction Schemes: single substrate multiple substrate single displacement double disp (ping-pong)
 noncompetitive (rev) mixed (rev) irreversible. Reaction Schemes: single substrate. multiple substrate. single displacement. double disp (ping-pong)")
19
Reaction Rate vs. Enzyme and Substrate Conc.

20
Control of Enzyme Activity
We don’t always need high levels of products of enzyme-catalyzed reactions around. What kind of control system is used to regulate amounts of enzyme and products? Two main methods: zymogens, and feedback control.

21
Zymogens Many enzymes are active as soon as they’re made.
However, some are made in an inactive form and stored. This inactive form is called a zymogen or proenzyme. To become active, the body needs only to cleave off a small peptide fragment.

22
Feedback Control Some enzymes (allosteric enzymes) bind molecules called regulators (different from the substrate) that can affect the enzyme either positively or negatively Positive regulator: speeds up the reaction by changing the shape of the active site -- substrate binds more effectively Negative regulator: slows down reaction by preventing proper substrate binding, again, by changing enzyme shape Feedback control: the end product acts as a negative regulator. If there is enough of the end product, it will slow down the first enzyme in a pathway.
 bind molecules called regulators (different from the substrate) that can affect the enzyme either positively or negatively. Positive regulator: speeds up the reaction by changing the shape of the active site -- substrate binds more effectively. Negative regulator: slows down reaction by preventing proper substrate binding, again, by changing enzyme shape. Feedback control: the end product acts as a negative regulator. If there is enough of the end product, it will slow down the first enzyme in a pathway.")
23
The kinetics of enzyme catalysis: Steady state kinetics

24
A hyperbolic curve between V0 and [S] was revealed by in vitro studies using purified enzymes
It was the initial velocity (rate), V0, that was measured, so the change of [S] could be ignored. The catalysis was assumed to occur as: The enzyme will become saturated at high [S]: the V0 will not be affected by [S] at high [S].
![A hyperbolic curve between V0 and [S] was revealed by in vitro studies using purified enzymes](http://slideplayer.info/slide/4884800/16/images/24/A+hyperbolic+curve+between+V0+and+%5BS%5D+was+revealed+by+in+vitro+studies+using+purified+enzymes.jpg "It was the initial velocity (rate), V0, that was measured, so the change of [S] could be ignored. The catalysis was assumed to occur as: The enzyme will become saturated at high [S]: the V0 will not be affected by [S] at high [S].")
25
The effect on V0 of varying [S] is measured when the enzyme
Vmax is extrapolated from the plot: V0 approaches but never quite reaches Vmax. The effect on V0 of varying [S] is measured when the enzyme concentration is held constant. Hyperbolic relationship between V0 and [S]
![The effect on V0 of varying [S] is measured when the enzyme](http://slideplayer.info/slide/4884800/16/images/25/The+effect+on+V0+of+varying+%5BS%5D+is+measured+when+the+enzyme.jpg "Vmax is extrapolated from the plot: V0 approaches but never quite reaches Vmax. The effect on V0 of varying [S] is. measured when the enzyme. concentration is held constant. Hyperbolic relationship between. V0 and [S]")
26
A mathematical relationship between V0 and [S] was established (Michaelis and Menten, 1913; Briggs and Haldane, 1925) k 1 k 2 ( ) E + S ES E + P Formation of ES is fast and reversible. The reverse reaction from PS (k-2 step) was assumed to be negligible. The breakdown of ES to product and free enzyme is the rate limiting step for the overall reaction. ES was assumed to be at a steady state: its concentration remains constant over time. Thus V0 = k2[ES]
![A mathematical relationship between V0 and [S] was established (Michaelis and Menten, 1913; Briggs and Haldane, 1925)](http://slideplayer.info/slide/4884800/16/images/26/A+mathematical+relationship+between+V0+and+%5BS%5D+was+established+%28Michaelis+and+Menten%2C+1913%3B+Briggs+and+Haldane%2C+1925%29.jpg "k 1. k 2. ( ) E + S ES E + P. Formation of ES is fast and reversible. The reverse reaction from PS (k-2 step) was assumed to be negligible. The breakdown of ES to product and free enzyme is the rate limiting step for the overall reaction. ES was assumed to be at a steady state: its concentration remains constant over time. Thus V0 = k2[ES]")
27
k1([Et]-[ES])[S]=k-1[ES] + k2[ES]
Steady-state assumption: Rate of ES formation=rate of ES breakdown k1([Et]-[ES])[S]=k-1[ES] + k2[ES] ([Et] is the total enzyme concentration.) Solve the equation for [ES]: Km is called the Michaelis constant. V0 = k2[ES]
![k1([Et]-[ES])[S]=k-1[ES] + k2[ES]](http://slideplayer.info/slide/4884800/16/images/27/k1%28%5BEt%5D-%5BES%5D%29%5BS%5D%3Dk-1%5BES%5D+%2B+k2%5BES%5D.jpg "Steady-state assumption: Rate of ES formation=rate of ES breakdown. k1([Et]-[ES])[S]=k-1[ES] + k2[ES] ([Et] is the total enzyme concentration.) Solve the equation for [ES]: Km is called the. Michaelis constant. V0 = k2[ES]")
28
The maximum velocity is achieved when all the enzyme is saturated by substrate, i.e., when
[ES] =[Et]. Thus Vmax =k2[Et] The Michaelis-Menten Equation

29
The Michaelis-Menten Equation nicely
When [S] >> Km When [S] << Km The Michaelis-Menten Equation nicely describes the experimental observations. The substrate concentration at which V0 is half maximal is Km

30
The Vmax and Km values of a certain enzyme can be measured by the double reciprocal plot (i.e., the Lineweaver-Burk plot).
.")
31
The double reciprocal plot: 1/V0 vs 1/[S]
![The double reciprocal plot: 1/V0 vs 1/[S]](http://slideplayer.info/slide/4884800/16/images/31/The+double+reciprocal+plot%3A+1%2FV0+vs+1%2F%5BS%5D.jpg "The double reciprocal plot: 1/V0 vs 1/[S]")
32
The Michaelis-Menten equation, but not their approximated mechanism applies to a great many enzymes
Most enzymes (except the regulatory enzymes) have been found to follow the Michaelis-Menten kinetics, but their actual mechanisms are usually more complicated (by having more intermediate steps) than the one assumed by Michaelis and Menten. The values of Vmax and Km alone provide little information about the number, rates, or chemical nature of discrete steps in the reaction.
 have been found to follow the Michaelis-Menten kinetics, but their actual mechanisms are usually more complicated (by having more intermediate steps) than the one assumed by Michaelis and Menten. The values of Vmax and Km alone provide little information about the number, rates, or chemical nature of discrete steps in the reaction.")
33
The actual meaning of Km depends on the reaction mechanism
For If k2 is rate-limiting, k2<<k-1, Km = k-1/k 1 Km equals to the dissociation constant (Kd) of the ES complex; Km represent a measure of affinity of the enzyme for its substrate in the ES complex.
 of the ES complex; Km represent a measure of affinity of the enzyme for its substrate in the ES complex.")
35
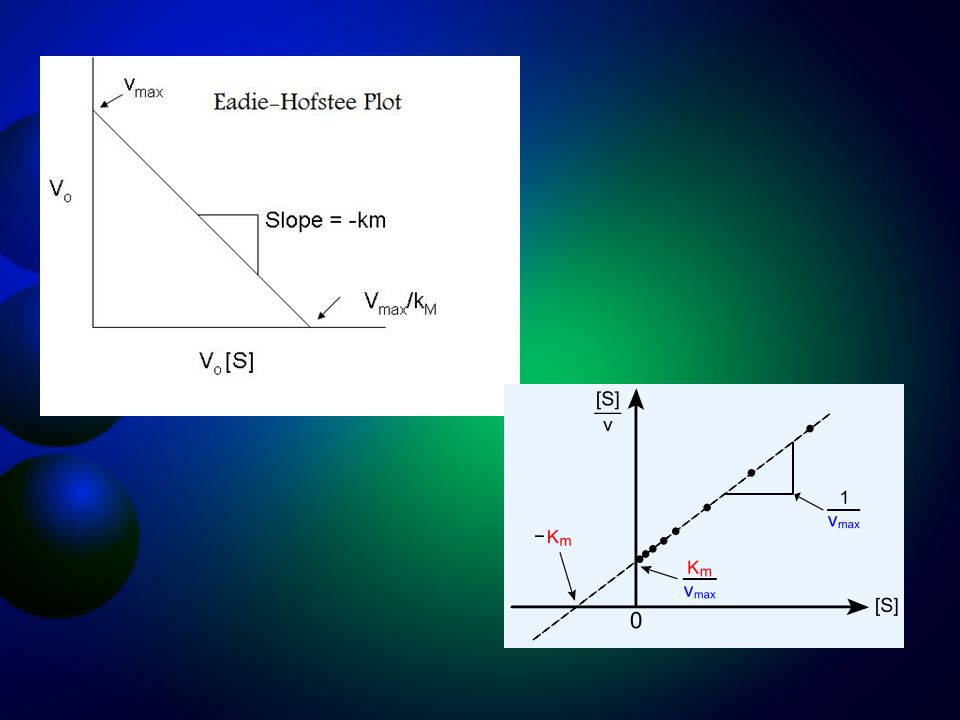
PLOT EADIE-HOFSTEE DAN HANES - WOOLF
Plot Lineweaver – Burk mempunyai sedikit kelemahan, yaitu Sering kali pada saat mengekstrapolasi grafik untuk menentukan harga -1/Km ternyata akan memotong sumbu 1/[S] di luar grafik yang dibuat Pada konsentrasi substrat yang terlalu rendah, maka akan diperoleh hasil yang kurang akurat Awal dari kelinearannya sering kurang jelas dibanding dengan plot lain, terutama plot Eadie – Hofstee, padahal hal ini sangat penting pada penentuan mekanisme reaksi

36
Plot Eadie-Hofstee dan Hanes diturunkan dari persamaan Lineweaver-Burk dengan mengalikan kedua sisi persamaan dengan faktor vo Vmax sehingga akan diperoleh persamaan garis lurus selanjutnya dipergunakan untuk menghitung Vmax dan Km

37
Dengan cara penurunan yang mirip, Hanes-Woolf mengalikan perasamaan Lineweaver-Burk dengan [So] maka diperoleh: Plot Eadie – Hofstee dan Hanes banyak digunakan pada studi kinetik enzim, namun demikian studi enzim secara umum masih menggunakan plot Lineweaver – Burk.
![Dengan cara penurunan yang mirip, Hanes-Woolf mengalikan perasamaan Lineweaver-Burk dengan [So] maka diperoleh:](http://slideplayer.info/slide/4884800/16/images/37/Dengan+cara+penurunan+yang+mirip%2C+Hanes-Woolf+mengalikan+perasamaan+Lineweaver-Burk+dengan+%5BSo%5D+maka+diperoleh%3A.jpg "Plot Eadie – Hofstee dan Hanes banyak digunakan pada studi kinetik enzim, namun demikian studi enzim secara umum masih menggunakan plot Lineweaver – Burk.")
39
Lineweaver-Burk (double reciprocal plot)
Rewrite Michaelis-Menten rate expression Plot 1/v versus 1/[S]. Slope is Km/Vmax, intercept is 1/Vmax

40
Graphical Solution intercepts 1/ V Slope = Km/ Vmax 1/ Vmax -1/ Km

41
Example: Lineweaver-Burk

42
Resulting Plot slope = Km/ Vmax= y intercept = 1/ Vmax=

43
Michaelis-Menten Kinetics

44
Vmax = 1/ x 10-4 = 3.49 x 10-5 M/min Km= x Vm = 1.98 x 10-5 M

45
Other Methods Eadie-Hofstee plot Hanes- Woolf

46
Comparison of Methods Lineweaver-Burk: supposedly gives good estimate for Vmax, error is not symmetric about data points, low [S] values get more weight Eadie-Hofstee: less bias at low [S] Hanes-Woolf: more accurate for Vmax. When trying to fit whole cell data – I don’t have much luck with any of them!

47
PERSAMAAN HALDAN UNTUK REAKSI REVERSIBEL
Reaksi enzimatis dalam sel sering berlangsung secara reversibel. Reaksi substrat tunggal, S P, berlangsung melalui pembentukan satu kompleks intermediate, arah ke kanan dianggap sebagai kompleks ES dan sebaliknya kompleks EP E + S ES/EP P + E

48
Persamaan MM arah kekanan pada [Eo] tetap dengan laju awal vf dan Vsmax
Persamaan MM ke arah sebaliknya pada [Eo] tetap dengan laju awal vb dan VPmax
![Persamaan MM arah kekanan pada [Eo] tetap dengan laju awal vf dan Vsmax](http://slideplayer.info/slide/4884800/16/images/48/Persamaan+MM+arah+kekanan+pada+%5BEo%5D+tetap+dengan+laju+awal+vf+dan+Vsmax.jpg "Persamaan MM ke arah sebaliknya pada [Eo] tetap dengan laju awal vb dan VPmax.")
49
Perumusan Haldan hubungan antara konstanta laju dan kesetimbangan reaksi pada reaksi kesetimbangan adalah Karena

50
Maka: Bila konstanta kesetimbangan diketahui, maka persamaan tersebut dapat digunakan untuk memvalidasi konstanta laju yang diperoleh Secara umum Km dari arah reaksi metabolisme penting akan sedikit lebih kecil dari arah sebaliknya. Namun arah metabolisme dipengaruhi juga oleh [S] dan [P] dalam sel

51
KINETIKA REAKSI CEPAT Kinetika keadaan steady penting bagi enzimologis yang memungkinkan untuk menentukan Km dan Kcat Bilangan peredaran (turn over number), Kcat pada beberapa enzim pada tingkat 100 s-1, yaitu 100 molekul produk dihasilkan setiap detik tiap molekul enzim. Hal ini berarti bahwa tahapan yang paling lambat dari suatu mekanisme reaksi mempunyai waktu hidup hanya beberapa detik
, Kcat pada beberapa enzim pada tingkat 100 s-1, yaitu 100 molekul produk dihasilkan setiap detik tiap molekul enzim. Hal ini berarti bahwa tahapan yang paling lambat dari suatu mekanisme reaksi mempunyai waktu hidup hanya beberapa detik.")
52
Untuk reaksi sederhana order pertama
A B Maka laju reaksi pada t adalah -d[A]/dt = d[B]/dt = k[A] Integrasi dari persamaan tersebut memberikan persamaan ln[Ao] – ln [A] = kt [A] = [Ao]e –kt

53
konsentrasi A B waktu Kurva teoritis perubahan reaksi A menjadi B

54
Persamaan integrasi dari sebagian besar mekanisme reaksi akan lebih rumit . Untuk reaksi substrat tunggal yang membentuk suatu kompleks intermediate dengan konsentrasi awal dari [S] >>> [E] dinyatakan sebagai: E + S ES E + P Laju penambahan [ES] pada waktu t (pada periode awal dimana pembentukan [P] diabaikan dinyatakan sebagai: d[S]/dt = [E][S] [ES] [ES] d[S]/dt = ([Eo] - [ES])([So] – [ES] – [P]) [ES] [ES] karena : [So]>>[Eo], maka ([So]-[ES]-[P] ~ [So] Jadi : d[S]/dt = ([Eo] - [ES])([So] ) [ES] [ES] k2 k1 k-1 k-2 k1 k-1 k2 k1 k-1 k2 k1 k-1 k2
![Persamaan integrasi dari sebagian besar mekanisme reaksi akan lebih rumit . Untuk reaksi substrat tunggal yang membentuk suatu kompleks intermediate dengan konsentrasi awal dari [S] >>> [E] dinyatakan sebagai:](http://slideplayer.info/slide/4884800/16/images/54/Persamaan+integrasi+dari+sebagian+besar+mekanisme+reaksi+akan+lebih+rumit+.+Untuk+reaksi+substrat+tunggal+yang+membentuk+suatu+kompleks+intermediate+dengan+konsentrasi+awal+dari+%5BS%5D+%3E%3E%3E+%5BE%5D+dinyatakan+sebagai%3A.jpg "E + S ES E + P. Laju penambahan [ES] pada waktu t (pada periode awal dimana pembentukan [P] diabaikan dinyatakan sebagai: d[S]/dt = [E][S] - [ES] - [ES] d[S]/dt = ([Eo] - [ES])([So] – [ES] – [P]) - [ES] - [ES] karena : [So]>>[Eo], maka ([So]-[ES]-[P] ~ [So] Jadi : d[S]/dt = ([Eo] - [ES])([So] ) - [ES] - [ES] k2. k1. k-1. k-2. k1. k-1. k2. k1. k-1. k2. k1. k-1. k2.")
55
Integrasi persamaan tersebut akan menunjukkan perubahan ES terhadap waktu
E + S ES EP E + P dimana [So] >> [Eo] Konstr. S P E ES Induction period Steady state phase Waktu Transient or pre steady state phase

56
Bagian linier dari grafik [P] vs t menunjukkan fase steady state dari reaksi dengan slope = k2[E][So]/([So]/Km) yang didapat dari mensubstitusi (k-1 + k2)/k1 dengan Km dari persamaan integrasinya Jika bagian grafik steady state linier, ekstrapolasinya akan memotong sumbu t pada t = 1/(k1[So]+Km) dan disebut sebagai periode induksi. Kurva perubahan konsentrasi akan lebih rumit bila reaksinya seperti; Dengan tahap laju yang menentukan (rate-limiting-step) adalah EP menjadi E dan P. E + S ES EP E + P
![Bagian linier dari grafik [P] vs t menunjukkan fase steady state dari reaksi dengan slope = k2[E][So]/([So]/Km) yang didapat dari mensubstitusi (k-1 + k2)/k1 dengan Km dari persamaan integrasinya](http://slideplayer.info/slide/4884800/16/images/56/Bagian+linier+dari+grafik+%5BP%5D+vs+t+menunjukkan+fase+steady+state+dari+reaksi+dengan+slope+%3D+k2%5BE%5D%5BSo%5D%2F%28%5BSo%5D%2FKm%29+yang+didapat+dari+mensubstitusi+%28k-1+%2B+k2%29%2Fk1+dengan+Km+dari+persamaan+integrasinya.jpg "Jika bagian grafik steady state linier, ekstrapolasinya akan memotong sumbu t pada t = 1/(k1[So]+Km) dan disebut sebagai periode induksi. Kurva perubahan konsentrasi akan lebih rumit bila reaksinya seperti; Dengan tahap laju yang menentukan (rate-limiting-step) adalah EP menjadi E dan P. E + S ES EP E + P.")
57
E + S ES EP E + P dimana EP menjadi E dan P sebagai penentu
Konstr. EP E P ES Waktu

58
Vmax is determined by kcat, the rate constant of the rate-limiting step
Vmax = kcat[Et] kcat equals to k2 or k3 or a complex function of both, depending on which is the rate-limiting step. kcat is also called the turnover number: the number of substrate molecules converted to product in a given unit of time per enzyme molecule when the enzyme is saturated with substrate.

59
40,000,000 molecules of H2O2 are converted
to H2O and O2 by one catalase molecule within one second!

60
The kinetic parameters kcat and Km are often studied and compared for different enzymes
Km often reflects the normal substrate concentration present in vivo for a certain enzyme. The catalytic efficiency of different enzymes is often compared by comparing their kcat/Km ratios (the specificity constant). kcat/Km is an apparent second-order rate constant (with units of M-1S-1), relating the reaction rate to the concentrations of free enzyme and substrate.
. kcat/Km is an apparent second-order rate constant (with units of M-1S-1), relating the reaction rate to the concentrations of free enzyme and substrate.")
61
The value of kcat/Km has an upper limit (for the perfected enzymes)
It can be no greater than k1. The decomposition of ES to E + P can occur no more frequently that E and S come together to form ES. The most efficient enzymes have kcat/Km values near the diffusion-controlled limit of 108 to 109 M-1S-1.

62
Catalytic perfection (rate of reaction being
diffusion-controlled) can be achieved by a combination of different values of kcat and Km.
 can be achieved by a. combination of different values of kcat and Km.")
63
Rate enhancement is often used to describe the efficiency of an enzyme
kcat catalyzed uncatalyzed kcat Rate enhancement: ratio of the rates of the catalyzed and the uncatalyzed reactions.

64
Rate enhancement by selected enzymes
Uncatalyzed rate (kun, s-1) Catalyzed rate (kcat, s-1) Rate enhancement (kcat/kun) Nonenzymatic half-life Enzyme
 Catalyzed. rate. (kcat, s-1) Rate. enhancement. (kcat/kun) Nonenzymatic. half-life. Enzyme.")
65
Enzyme-catalyzed reactions of two or more substrates can also be analyzed by the Michaelis-Menten approach Each substrate will have one characteristic Km value. Noncovalent ternary complex (with two substrates bound to the enzyme concurrently) may or may not be formed for the bisubstrate reactions depending on the mechanism. Steady-state kinetics can often help distinguish these two mechanisms.
 may or may not be formed for the bisubstrate reactions depending on the mechanism. Steady-state kinetics can often help distinguish these two mechanisms.")
66
In those enzyme-catalyzed bisubstrate reactions where a
ternary complex is formed, the two substrates may either bind in a random sequence or in a specific order.

67
For those reactions where ternary complex is formed:
Maintaining the concentration of one substrate (S2) constant, the double reciprocal plots made by varying the concentration of the other substrate (S1) will intersect.
 constant, the double reciprocal plots. made by varying the concentration. of the other substrate (S1) will. intersect.")
68
No ternary complex is formed in the Ping-Pong (or double displacement) mechanism: The first substrate is converted to a product that leaves the enzyme active site before the second substrate enters.
 mechanism: The first substrate is converted to a product that leaves the enzyme active site before the second substrate enters.")
69
As [S2] increases, Vmax increases,
For enzymes having Ping-Pong mechanisms (ternary complex not formed). Maintaining the concentration of one substrate (S2) constant, the double reciprocal plots made by varying the concentration of the other substrate (S1) will not intersect. As [S2] increases, Vmax increases, as does the Km for S1. S1
![As [S2] increases, Vmax increases,](http://slideplayer.info/slide/4884800/16/images/69/As+%5BS2%5D+increases%2C+Vmax+increases%2C.jpg "For enzymes having Ping-Pong mechanisms (ternary. complex not formed). Maintaining the concentration. of one substrate (S2) constant, the double reciprocal plots. made by varying the concentration. of the other substrate (S1) will not. intersect. As [S2] increases, Vmax increases, as does the Km for S1. S1.")
70
Rates of individual steps for an enzyme-catalyzed reaction may be obtained by pre-steady state kinetics The enzyme (of large amount) is used in substrate quantities and the events on the enzyme are directly observed. Rates of many reaction steps may be measured independently. Very rapid mixing and sampling techniques are required (the enzyme and substrate have to be brought together in milliseconds and measurements also be made within short period of time).
 is used in substrate quantities and the events on the enzyme are directly observed. Rates of many reaction steps may be measured independently. Very rapid mixing and sampling techniques are required (the enzyme and substrate have to be brought together in milliseconds and measurements also be made within short period of time).")
71
“Rapid kinetics” or “pre-steady- state kinetics”is applied to the
observation of rates of systems that occur in very short time intervals (usually ms or sub-ms scale ) and very low product concentrations. This period covers the time from the enzyme encountering its target (either a substrate, inhibitor or some other ligands) to the point of system settling to equilibrium. The concentration of ES will rise from zero to its steady-state value. (ms or sub-ms)
 and very low product. concentrations. This period. covers the time from the enzyme. encountering its target (either. a substrate, inhibitor or some. other ligands) to the point of. system settling to equilibrium. The concentration of ES will rise. from zero to its steady-state value. (ms or sub-ms)")
72
The turnover number of carbonic anhydrase:
Carbonic anhydrase of erythrocytes (Mr 30,000) has one of the highest turnover numbers among known enzymes, it catalyses the reversible reaction of CO2: H2O + CO2 -> H2CO3 This is an important process in the transport of CO2 from the tissues to the lungs. If 10μg of pure carbonic anhydrase catalyses the hydration of 0.30g of CO2 in 1min at 37°C at Vmax, and the reaction volume is 1ml. What is the turnover number (Kcat) of carbonic anhydrase expressed in units of per min and per sec)? Mr of CO2 is 44.
 has one of the highest turnover numbers among known enzymes, it catalyses the reversible reaction of CO2: H2O + CO2 -> H2CO3. This is an important process in the transport of CO2 from the tissues to the lungs. If 10μg of pure carbonic anhydrase catalyses the hydration of 0.30g of CO2 in 1min at 37°C at Vmax, and the reaction volume is 1ml. What is the turnover number (Kcat) of carbonic anhydrase expressed in units of per min and per sec) Mr of CO2 is 44.")
Presentasi serupa















>")




